КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Механика мышечного сокращения
МЕХАНИКА ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
Основными материальными структурами опорно-двигательного аппарата являются кости скелета, мышцы, а также связки и сухожилия, объединяющие все компоненты в единую систему. Скелет выполняет опорную функцию и обеспечивает сохранение геометрической формы организма. Большинство костей скелета контактируют друг с другом при помощи подвижных сочленений (суставов), устройство которых определяет характер возможных движений. В качестве силового элемента служат мышцы, в которых для генерации механических сил и совершения работы используется энергия, запасенная в АТФ. Величины, направления сил, характер движения костей скелета, количество выполненной работы должны зависеть от структуры, физических и биологических свойств мышц, а также их расположения в пространстве.
По современным представлениям все мышцы разделяются на три группы: 1. скелетные, 2. гладкие, 3. мышца сердца (миокард). Миокард представляет собой основную массу сердца. При его сокращении уменьшается объем полостей сердца (предсердий, желудочков), повышается давление, которое обеспечивает непрерывное течение крови.
Гладкие мышцы входят в состав стенок внутренних органов (желудка, кишечника, сосудов, мочевого пузыря и т.п.). При напряжении этих мышц происходит изменение формы органов, необходимое для осуществления биологических процессов (движения крови, лимфы, мочи, пищи и др.).
|

Скелетные мышцы состоят из большого количества параллельно расположенных волокон, представляющих собой высокоспециализированные клетки. Как и любые другие клетки, они отграничены мембраной и содержат органоиды: ядра, саркоплазматический ретикулум, митохондрии и др. Особое значение для выполнения сократительной функции принадлежит тонким (диаметром 1-2 мкм) нитям - миофибриллам, которые тянутся от одного конца волокна до другого. В первом приближении можно считать, что длина миофибрилл равна длине мышцы. Количество этих органоидов в клетке достаточно велико и может достигать 1000-2000.
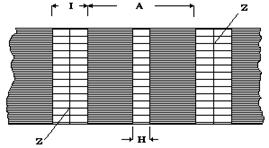
|
C помощью микроскопических наблюдений в миофибриллах выявляются поперечно расположенные зоны А, I, Н, Z (см. рис. 10). Наличие поперечной исчерченности дало основание называть скелетные мышцы поперечно-полосатыми. Отметим, что в мышце сердца также обнаруживается поперечная исчерченность.
В миофибриллах можно выделить последовательно
|
Рис. 10 повторяющийся элемент - саркомер, ограниченный Z - дисками, исходная длина которого уменьшается в процессе сокращения. По современным представлениям именно саркомер является элементарной моторной единицей, а механизм его сокращения лежит в основе деятельности всех скелетных мышцы. При уменьшении длины саркомера (расстояния между Z-дисками)
Рис. 11 изменяются размеры и взаиморасполо-жение зон А, I, H. Как показывают результаты электронно-микроскопического наблюдения, эти сдвиги объясняются взаимодействием более тонких нитей - протофибрилл (см. рис.11).
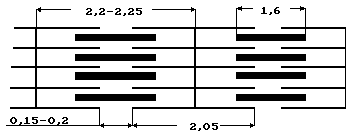
Выявлены протофибриллы двух типов - тонкие, состоящие из белка актина, и толстые, содержащие белок - миозин. Сопоставление результатов электронной и оптической микроскопии позволяет объяснить поперечную исчерченность: Н - расстояние между концами актиновых нитей в центре саркомера, I - промежуток между концами миозиновых нитей рядом расположенных саркомеров. В основе теоретического объяснения сокращения саркомера лежит модель скользящих нитей.
|
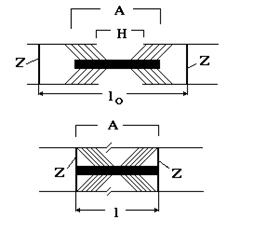
При раздражении мышц нервными импульсами или электрическим током из саркоплазматического ретикулума освобождаются ионы кальция, которые способствуют формированию связей (мостиков) между миозиновыми и актиновыми нитями (см. рис. 12). Образующийся акто-миозиновый комплекс обладает ферментативной активностью и расщепляет универсальный источник энергии - молекулы АТФ. Освобожденная при этом энергия передается на кальциевые мостики, которые работают в циклическом режиме. Мостики обеспе-
Рис. 12 чивают движение актиновых нитей к центру полосы в Н-зону в результате чего Z-перегородки приближаются к концам миозиновых нитей. Этот молекулярный механизм создает усилие, способствующее перемещению протофибрилл мышечного волокна друг относительно друга до тех пор, пока концы миозиновых нитей не достигнут Z-дисков (см. рис. 12).
По существующим оценкам при расщеплении молекулы АТФ один мостик за полный цикл работы в течение 1 мс развивает усилие 10-12 H. Поскольку саркомеры располагаются в миофибрилле последовательно, исходную длину мышцы, можно выразить Lo = nlo, где lo - длина саркомера до сокращения, n - количество саркомеров. В процессе уменьшения расстояния между Z - дисками на Dl = lo - l длина всей миофибриллы изменяется на DL = Lo - L = nlo - nl = n(lo - l) = n Dl. Отношение величины укорочения мышцы к ее исходной длине Lo называется относительным укорочением: e = DL/Lo = n Dl/nlo = Dl /lo. Как видно из приведенного соотношения, относительное укорочение мышцы можно рассчитать следующим образом. Воспользовавшись геометрическими размерами, определенными из микроскопических наблюдений (длина lo = 2.00 - 2.25 мкм, размер зоны А a = 1.6 мкм), можно определить абсолютное Dl = lo - a = 0.6 - 0.65 мкм и относительное укорочение саркомера e= Dl/lo =0.27 - 0.29 = 27 - 29%.
Аналогичные расчеты показывают, что максимальное относительное укорочение для большинства мышц составляет величину порядка 30%. Следовательно, для получения зна-чительных перемещений DL = eLo необходимо, чтобы движение обеспечивалось мышцами с большой исходной длиной.
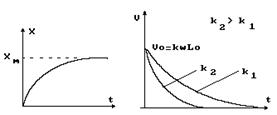
Определим зависимость изменения длины мышцы от времени. Очевидно, скорость сокращения в начальный момент времени наибольшая, затем уменьшается и, когда укорочение достигает максимума DL, становится равной нулю. Такая зависимость может быть описана следующим выражением: V = k(DL - X), где Х - укорочение в момент времени DL - наибольшее укорочение при t ® ¥ и k - коэффициент пропорциональности, зависящий от приложенной к мышце нагрузки. Скорость сокращения является первой производной от укорочения повремени: V = dX/dt = k(DL - X). Решая это дифференциальное уравнение, найдем выражение, описывающее укорочение во времени: X = DL (1 - e- k t ) или X = eLo (1 - e- k t ), где e - относительное укорочение, Lo - исходная длина мышцы. Скорость сокращения определится как первая производная от укорочения:
V = dX/dt = ke Lo e-k t = Vo e-k t
На рисунке 13 приведены графики зависимости укорочения Х и скорости V от времени t.
|
Если миофибриллы располагаются вдоль оси мышцы, количество мостиков между протофибриллами зависит от площади поперечного сечения мышцы S, и поэтому силу сокращения можно выразить: F = = f ´ S, где S- площадь сечения, f - удельная сила сокращения, которую развивают мостики на единице сечения. Последняя величина для скелетных
Рис. 13 мышц человека составляет 30 - 40 Н/см2.
|
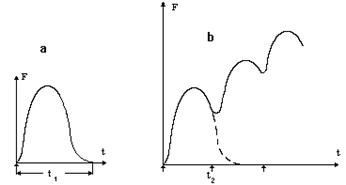
В зависимости от условий стимуляции наблюдается различный характер деятельности мышц: одиночное сокращение, зубчатый и гладкий тетанус. Когда на мышцу воздействует одиночный электрический импульс, сила, которую она развивает, изменяется во времени по закону, приведенному на рисунке 14а. Такое сокращение называется одиночным и заканчивается в момент t1.
Если воздействовать серией электрических раздражителей с частотой f = 1/t2 причем t2 < t1, то новый электрический
Рис. 14 импульс будет раздражать мышцу в момент, когда она еще полностью не расслабилась и не восстановила исходную длину. В этом случае развиваемая сила не достигает нуля, а изменяется по закону, приведенному на рисунке 14b. Такой режим сокращения называется зубчатым тетанусом.
|
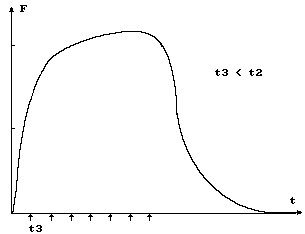
Если раздражать мышцу с еще большей частотой f = 1 / t3, она вообще не успевает расслабляться и создаваемая сила описывается гладкой кривой. Подобное сокращение называется гладким тетанусом (см. рис. 15). При гладком тетанусе напряжение мышцы превышает напряжение одиночного сокращения в 1,5 - 2 раза.
Это связано с тем, что в мышце имеются
упругие элементы (внефибриллярные компо-
Рис. 15 ненты соединительной ткани), которые при активном укорочении мышцы скручиваются наподобие пружины и препятствуют развитию напряжения в сократительных элементах за время t1. При переходе одиночных сокращений в тетанус последовательно расположенные упругие элементы деформируются и не оказывают такого противодействия, как при одиночном сокращении. Необходимо отметить, что этот механизм играет решающую роль в расслаблении мышц. После сокращения ионы кальция удаляются из цитоплазмы в саркоплазматический ретикулум, актомиозиновый комплекс распадается и за счет внутреннего напряжения деформированных упругих элементов протофибриллы возвращаются в исходное состояние. Величину механической работы в процессе мышечного сокращения можно выразить: А = F ´ DL = f S DL, где F мышечная сила (при условии, что она постоянная), DL - абсолютное укорочение, равное L = Lo - L.
Считая в первом приближении, что мышца может быть представлена цилиндром, умножим и поделим величину работы на исходную длину Lo:
А = f S DL 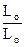
Произведение S Lo выражает исходный объем Vo, а e = DL/Lo относительное укорочение мышцы. Тогда работа мышцы А = f e Vo. Если представить объем через плотность и массу мышцы Vo = M/r, то:
А = e M (f / r)
Отношение удельной силы к плотности для данной мышечной ткани является постоянной величиной k = f / r, и тогда окончательно работа при мышечном сокращении:
А = k e M.
|
Из этого равенства следует, что механическая работа зависитот массы и относительного укорочения мышцы. Величина силы, которую развивают протофибриллы, определяется длиной саркомера. Это объясняется тем, что в процессе скольжения нитей, во-первых, изменяется количество мостиков, образующихся между протофибриллами, а во-вторых, -
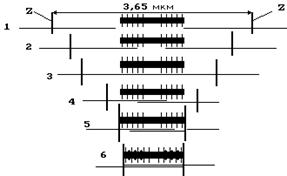
Рис. 16 при минимальной длине саркомера миозиновые нити упираются в Z -перегородки (см. рис. 16). На рисунке 17 приведена экспериментальная зависимость между относительной силой и длиной саркомера. F% = (F/Fm)´100 (где F - развиваемая сила, а Fm - максимальная сила сокращения). При длине саркомера, l > 3.65 мкм между протофибриллами не могут образовываться мостики, и поэтому сила равна нулю. По мере уменьшения расстояния между активновыми и миозиновыми нитями образуется все большее количество мостиков и поэтому сила возрастает. Наибольшую силу саркомер развивает, когда l
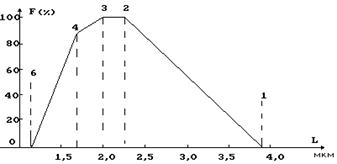
|
лежит в диапазоне 2 - 2,25 мкм. Если расстояние l становится меньше 2,05 мкм, актиновые нити перекрываются и усилие F падает.
При расстоянии между Z-дисками меньше 1,65 мкм концы миозиновых
нитей упираются в перегородки, они деформируются, нарушается их пространствен-
ная структура и поэтому развиваемая сила
Рис. 17 становится равной 0 (при l = 1,05 мкм).
|
Дата добавления: 2014-01-15; Просмотров: 1703; Нарушение авторских прав?; Мы поможем в написании вашей работы!