КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
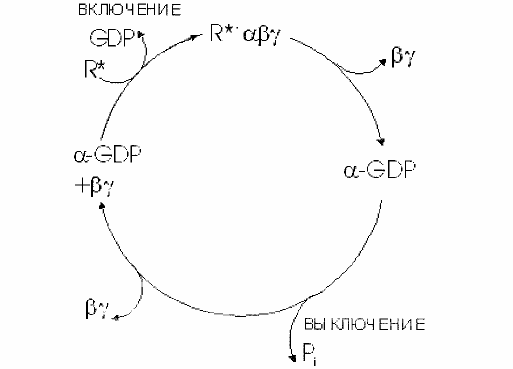
ГТФ-связывающие белки образуют два основных семейства G-белков и низкомолекулярных ГТФ-связывающих белков
|
|
|
|
Все эти белки сильно связывают ГТФ и превращают его в ГДФ, при этом происходит переход белка из активированного в неактивное состояние. Свойства ГТФ-связывающих белков. Основной структурной особенностью ГТФ-связывающих белков является наличие домена связывания гуаниновых нуклеотидов. Наличие соответствующих консенсусных аминокислотных последовательностей является однозначным указанием на принадлежность белка к данному семейству.
Литература
1. Биохимия.3-е издание (исправленное). М; Издательская группа ГЭОТАР-Медиа. 2006г.
2. Основы биохимии. А. Уайт, Ф Хендлер, Э. Смит, Р. Хилл, И. Леман.3 т. М; Мир1981 г.
3. Биохимия. Марри, Греннер. М; Высшая школа. 1993г.
4. Биохимия. Ленинджер, 1995г.
5. Биохимия. В.П. Комов., В.Н. Шведова. Дрофа; М: 2004г..
6. Общая биохимия. Учебное пособие по биохимии. Курс лекций. М.Т. Генгин; 1997г.
7. Основы биохимии. Ю.Б. Филиппович. Издание второе, переработанное и дополненное. Высшая школа; М4, 1995г.
Регуляция активности мембраносвязанных аденилатциклаз млекопитающих на уровне цитоплазматических доменов
Регуляция субъединицами Gs и Gi белков. Gs и Gi подсемейства G белков выступают в качестве сопрягающего звена между активированным рецептором и аденилатциклазой, ответственной за изменение концентрации цАМФ в клетке. Гормональные рецепторы, связанные с Gs и Gi подсемействами G белков, могут изменять аденилатциклазную активность через взаимодействие высвобождаемых a - и bg -субъединиц с циклазой. Изоформы мембраносвязанной аденилатциклазы млекопитающих по-разному отвечают на различные субъединицы G белков: одни и те же субъединицы G белка могут стимулировать некоторые изоформы, ингибировать другие и быть индифферентными к остальным изоформам. Все многообразие данных о регуляции мембраносвязанных аденилатциклаз млекопитающих субъединицами G белков, представленных в ряде фундаментальных обзоров [14,19,28], можно суммировать следующим образом.
|
|
|
1. Основной механизм активации мембраносвязанных аденилатциклаз млекопитающих осуществляется путем их взаимодействия с Gsa-субъединицей Gs белка. Все девять изоформ мембраносвязанной аденилатциклазы млекопитающих активируются ГТФ-связанной формой Gsa.
При Gsa стимуляции АЦ I, связанной с мембранами клеток насекомых (Sf9 или Hi-5), выявляется единственный сайт связывания. В то же время Gsa стимуляция АЦ II и АЦ VI (в тех же условиях) лучше объяснялась моделью двух не взаимодействующих между собой и сайтов связывания. Поскольку мембраносвязанные аденилатциклазы млекопитающих содержат только один сайт связывания Gsa, то для объяснения этих данных было выдвинуто предположение о возможной димеризации аденилатциклаз в мембране. Исходя из этого, можно полагать, что АЦ I имеет очень низкую способность к образованию гомодимеров, в то время как у АЦ II и АЦ VI эта способность резко выражена. Эти различия могут служить для объяснения разнообразных ответов различных клеток и тканей на гормоны, которые повышают уровень цАМФ.
2. Регуляция аденилатциклазной активности гормональными рецепторами, связанными с Gi- подсемейством G белков, является более сложной. Gia-субъединица вызывает заметное ингибирование Gsa- и форсколинстимулируемой активности АЦ V и АЦ VI. Gia-субъединица ингибирует также АЦ I. Следует отметить, что этот эффект отсутствовал в том случае, когда исследовалось Gsa-стимулирующее действие; ингибирование в значительной степени связано с активностью, наблюдаемой в присутствии кальмодулина или форсколина.
Показано, что Gia-субъединица связывается с АЦ V в полости, сформированной a2 и a3 спиралями С1 домена, которая является аналогичной для Gsa-связывающего сайта на С2 домене. Gia-субъединица уменьшает аффинность между С1 и С2 доменами и поэтому нарушает образование каталитического сайта аденилатциклазы. Мутанты, которые повышают аффинность С1 и С2 доменов друг к другу, понижают способность Gia-субъединицы к ингибированию фермента. Кроме того, Gia может оказывать влияние на связывание различных молекул с активным центром аденилатциклазы. Так, АЦ V может связывать аналоги АТФ в присутствии Gia, но не может одновременно связывать Gia и аналоги переходного состояния (2`a3`- АМФ). Gia не может также ингибировать АЦ V в присутствии ионов Mn2+, которые повышают аффинность аденилатциклазы к АТФ и аналогам субстрата.
|
|
|
Наряду с Gia, активность АЦ I, АЦ V и АЦ VI также ингибируется Gza-субъединицей. G0a-субъединица ингибирует только активность АЦ I. Активность АЦ II и АЦ III, стимулированная хорионическим гонадотропином в COS-7 клетках, также может ингибироваться Gia-субъединицей.
Доказано, что Gbg-комплекс непосредственно взаимодействует с АЦ I. С1a домен АЦ I (остатки 236-471) является достаточным для наблюдаемого bg-опосредованного ингибирования ферментативной активности, стимулированной Gsa или Са2+/кальмодулином. Интересно отметить, что в присутствии С1b области и С2 домена в молекуле АЦ I формируется второй сайт связывания Gbg. Ингибирование АЦ I может быть приписано взаимодействию bg-субъединиц с любым из этих двух сайтов. Для АЦ I, которая преимущественно экспрессируется в мозге, где G0 белок составляет около 1% от всех мембранных белков, активность фермента при стимуляции рецептора, связанного с G0 белком, преимущественно ингибируется Gbg-комплексом, а a-субъединица G0 белка способствует этому ингибированию.
Контрансфекция в клетках COS-7 АЦ V или АЦ VI с g2- и различными b-субъединицами приводит к значительному ингибированию активности обеих аденилатциклаз. Кроме того, показано, что b1 и b5 субъединицы различаются по ингибирующей активности. Так, Gb1 значительно ингибирует АЦ V и АЦ VI, в то время как Gb5 вызывает слабое ингибирование этих фермен.
В мозге и других тканях, которые экспрессируют АЦ II, АЦ IV и АЦ VII, в результате активации рецепторов, связанных с Gi и G0 белками, происходит высвобождение Gbg-комплексов, которые потенцируют Gsa-стимулируемую активность этих изоформ.
|
|
|
Сайт связывания в Gbg в АЦ II находится в области a3 спирали С2 домена. Вероятно, Gbg модулирует активность АЦ II, связываясь только с одним доменом (С2а), что приводит к такому изменению его конформации, которое вызывает оптимальное ориентационное упорядочение активного центра фермента. Сайт связывания Gbg в АЦ I отличается от такового в АЦ II, поскольку области a3-спиралей АЦ I и АЦ II значительно различаются между собой.
Gbg-комплексы не оказывают непосредственного влияния на ферментативную активность АЦ III, АЦ VIII и АЦ IX.
Регуляция ионами Са2+ и кальмодулином. Аденилатциклазная система и система обмена Са2+ - это две основные сигнальные системы, имеющиеся во всех клетках животных. Их функционирование взаимосвязано. Взаимодействие кальциевой и аденилатциклазной систем проявляется как на уровне регуляции или активности друг друга, так и посредством скоординированного влияния на биохимические процессы, происходящие в клетке.
Кальций-чувствительные аденилатциклазы, обеспечивают быстрый ответ цАМФ - генерирующей системы на повышение внутриклеточной концентрации Са2+ (положительный или отрицательный в зависимости от типа экспрессируемых аденилатциклаз).
Из девяти клонированных к настоящему времени изоформ аденилатциклаз млекопитающих пять изоформ оказались чувствительными к ионам Са2+ в условиях in vitro; АЦ I, АЦ III и АЦ VIII стимулируются Са2+, в то время как АЦ V и АЦ VI ингибируются Са2+ [19]. Многочисленные данные свидетельствуют о том, что эти изоформы фермента регулируются транзиторным изменением концентрации внутриклеточного Са2+ - [Ca2+]i. Так, в клетках НЕК 293, экспрессирующих АЦ I и АЦ VI, емкостный вход Са2+ (capacitative Ca2+ entry - CCE) вызывает стимуляцию и ингибирование, соответственно, синтеза цАМФ [7]. В клетках НЕК 293, экспрессирующих АЦ VIII, активация пуринергических рецепторов (которая повышает [Ca2+]i) приводит к стимуляции аденилатциклазной активности [30]. Было также показано, что в клетках С6 - 2В АЦ VI селективно ингибируется ССЕ в гораздо большей степени, чем Са2+, высвобождаемым из внутриклеточных депо [12]. Интересно отметить, что АЦ I и АЦ VIII, экспрессируемые в клетках НЕК 293, также заметно стимулируются ССЕ, в то вре
|
|
|
мя
как ионофоропосредованное высвобождение Са2+ из внутриклеточных депо не оказывает влияния на их активность [12].
АЦ III полностью рефрактерна к любому физиологическому повышению [Ca2+]i в клетках НЕК 293. При исследовании чувствительности к Са2+ этих аденилатциклаз в условиях in vitro была продемонстрирована значительная стимуляция АЦ I и АЦ VIII низкими микромолярными концентрациями Са2+. Наоборот,
АЦ III активируется только очень высокими супрамикромолярными концентрациями Са2+, которые являются ингибирующими для всех остальных аденилатциклаз млекопитающих [4]. Обнаружение того, что АЦ III рефрактерна к физиологически значимым подъемам концентрации внутриклеточного Са2+, а также ее активация чрезвычайно высокими концентрациями Са2+ в условиях in vitro предполагает ограниченную пригодность этой изоформы как интегратора кальциевых сигналов. Уникальность АЦ III заключается в том, что она активируется при тех концентрациях Са2+, при которых ингибируются все остальные изоформы аденилатциклазы [4]. Это свойство АЦ III может играть важную роль в тех случаях, когда синтез цАМФ должен осуществляться в присутствии высоких концентраций внутриклеточного Са2+. Предпочтение, которое отдают кальцийстимулируемые аденилатциклазы I и VIII емкостному входу Са2+ перед высвобождением Са2+ из внутриклеточных депо, предполагает дискретную локализацию аденилатциклаз в тех областях мембраны, где может поддерживаться достаточно высокий уровень [Ca2+]i [12]. Иными словами, предполагается колокализация Са2+ - чувствительных аденилатциклаз с каналами емкостного входа Са2+ в плазматической мембране клеток.
Это предположение находит подтверждение в иммунохимических исследованиях, которые указывают на то, что в нейронах аденилатциклазы сконцентрированы преимущественно в дендритических шипиках. Дендритические шипики являются областями с высокой концентрацией Са2+-каналов и насосов, а также заякоренных цАМФ-зависимых протеинкиназ. Поэтому дендритические шипики можно рассматривать как области с независимой нейрональной активностью и гомеостазом [Ca2+]i [13]. Все это выглядит таким образом, как если бы нейроны обладали способностью размещать аденилатциклазы там, где они являются больше всего подходящими для регуляции ионами Са2+ и эффективного распространения кальциевых сигналов. Такой характер локализации предполагает, что структура аденилатциклаз содержит информацию для их расположения в строго определенных микродоменах плазматической мембраны.
Перейдем к рассмотрению роли ионов Са2+ в регуляции Са2+-чувствительных аденилатциклаз в возбудимых тканях, в которых они преимущественно экспрессируются. Са2+-стимулируемые АЦ I и АЦ VIII экспрессируются исключительно в нейрональных клетках [30], в то время как Са2+-ингибируемые АЦ V и АЦ VI экспрессируются преимущественно в сердечной мышце [16]. Несмотря на значительное содержание в этих клетках потенциалзависимых кальциевых каналов (voltage-gated calcium channels-VGCCs), вызывает удивление полное отсутствие до последнего времени литературных данных о роли VGCC-опосредованного входа Са2+ на аденилатциклазную активность. Исключением было исследование, показывающее, что вход кальция через VGCCs ингибирует аденилатциклазную активность в вентрикулярных миоцитах цыпленка [34]. И только в 2000 году было показано, что гетерологически экспрессируемая АЦ VIII в клетках опухоли передней доли гипофиза линии GH4C1, стимулируется входом Са2+ через VGCCs L-типа [23]. В экспериментах с использованием нимодипина, который является эффективным блокатором VGCCs, оказалось, что емкостный вход кальция (ССЕ) вызывает эффективную стимуляцию АЦ VIII в клетках GH4C1, в то время как высвобождение кальция из внутриклеточных депо, вызываемое агонистами фосфолипазы С и ионофорами, не оказывало влияния на активность АЦ VIII [23]. Неэквивалентность в способности к подъему [Ca2+]i VGCC-опосредованного входа Са2+ (повышение Са2+ в цитозоле до 700 нм) и ССЕ-опосредованного входа Са2+ (повышение Са2+ в цитозоле до 250 нм) и равная эффективность активации АЦ VIII предполагает, что аденилатциклаза расположена гораздо ближе к ССЕ каналу, чем к VGCC в плазматической мембране.
Эти данные позволяют заключить, что близкие пространственные взаимоотношения в плазматической мембране между аденилатциклазой и ССЕ каналами, которые впервые были продемонстрированы в невозбудимых клетках, сохраняются и в возбудимых клетках, хотя аденилатциклаза является также чувствительной к значительному VGCC-опосредованному входу Са2+ в возбудимых клетках.
Полумаксимальное ингибирование нативной АЦ V из сердца собаки и ее растворимого гетеродимера VC1. VC2 наблюдается при концентрациях Са2+, соответственно, 7mМ и 5 mМ. В то же время ферментативная активность растворимого гетеродимера VC1a. VC2 не ингибируется Са2+ вплоть до концентрации 27 mМ. Было также показано, что 113-и аминокислотная С1b область (Arg 571- Phe 683) VC1 была способна связывать Са2+. Эти данные указывают на то, что С1b область АЦ V связывает Са2+ и делеция этой области в растворимом гетеродимере VC1. VC2 предотвращает ингибирование фермента ионами Са2+. Sholich и соавт. [8] полагают, что сайт связывания Са2+ в С1b области находится на участке Е 621 - D 652, в котором из 32 аминокислотных остатков 13 остатков носят кислотный характер. Большинство этих кислотных остатков являются высококонсервативными среди АЦ V и АЦ VI у животных разных видов. Поэтому предполагается, что эта 32-остаточная область в С1b является сайтом связывания Са2+ в АЦ V и АЦ VI. Интересно, отметить, что эквивалентным остатком Е 621 в АЦ V является D 602 в АЦ VI. Поскольку остатки аспарагиновой кислоты имеют более высокую аффинность для Са2+, чем остатки глутаминовой кислоты, возможно, что С1b область АЦ VI связывает Са2+ с большей аффинностью по сравнению с аналогичной областью в АЦ V. Это может объяснить большую чувствительность АЦ VI к ингибированию Са2+ по сравнению с АЦ V.
АЦ V и АЦ VI являются основными изоформами, экспрессируемыми в сердце [16]. Поскольку в течение сердечного цикла наблюдаются оcцилляции концентрации цАМФ [2], можно полагать, что ингибирование АЦ V и АЦ VI ионами Са2+ может служить в качестве отрицательной обратной связи, понижая уровень цАМФ в процессе сокращения.
Действительно, показано, что ионы Са2+ ингибируют катехолстимулируемое накопление цАМФ в кардиомиоцитах [34]. Экспериментальные данные предполагают, что Са2+-ингибируемые аденилатциклазы колокализованы с каналами емкостного входа Са2+ [6] и, поэтому, могут быть подвергнуты действию высоких концентраций свободного Са2+. Это согласуется с наблюдениями, что локальная концентрация свободного Са2+ в клетках может быть достаточно высокой, достигая значений 5 - 6 mМ [18]. Таким образом, АЦ V, которая полумаксимально ингибируется свободным Са2+ в концентрациях 5 - 7 mМ, может регулироваться путем изменения внутриклеточной концентрации Са2+.
Касаясь роли кальмодулина в регуляции Са2+-активируемых аденилатциклаз млекопитающих, следует отметить, что АЦ I,
АЦ III и АЦ VIII стимулируются Са2+ и кальмодулином в условиях in vitro. В то же время, в отличие от АЦ I и АЦ VIII, которые непосредственно стимулируются Са2+ и кальмодулином in vitro, АЦ III не стимулируется Са2+ и кальмодулином в отсутствие форсколина и a-субъединицы Gs белка [10]. Полумаксимальная стимуляция АЦ I и АЦ III наблюдается при концентрациях Са2+ 150 нМ и 5,0 mМ, соответственно. При исследовании регуляции АЦ III внутриклеточным Са2+ и рецепторами, связанными с Gs белком, в условиях in vivo оказалось, что гормон-стимулированная АЦ III ингибируется внутриклеточным Са2+. Механизм ингибирования АЦ III ионами Са2+ не зависит от активности цАМФ-зависимой протеинкиназы, протеинкиназы С или Gi белка. Поскольку ингибитор кальмодулин-зависимых киназ - KN 62 - блокирует этот эффект, можно полагать, что активированные Са2+ кальмодулин-зависимые киназы (КМ киназы) могут прямо или опосредованно ингибировать активность АЦ III in vivo. Действительно, коэкспрессия в клетках НЕК 293 АЦ III и активированной кальмодулин зависимой киназы II полностью ингибировала изопротеренол-стимулируемую активацию АЦ III [31].
Регуляция фосфорилированием/дефосфорилированием. Внутриклеточные концентрации цАМФ, достигнутые в ответ на экзогенные регуляторы, сильно зависят от состояния фосфорилирования/дефосфорилирования различных компонентов гормон-чувствительных аденилатциклазных систем (рецептор/
G белок/аденилатциклаза). Это особенно очевидно в случае рецепторов, которые десенситизируются путем фосфорилирования различными киназами: цАМФ-зависимыми киназами (ПКА), протеинкиназой С (ПКС) и рецептор-специфическими киназами, для которых субстратами являются агонистсвязанные формы рецепторов. Исследованию фосфорилирования различных субъединиц G белков также уделяется значительное внимание. В последние годы появились достаточно обширные данные о роли фосфорилирования/дефосфорилирования в регуляции аденилатциклаз.
Гормональная активация рецепторов, связанных с Gq и Gs белками, приводящая к активации ПКС и ПКА, соответственно, а также активация различных Са2+/кальмодулин-зависимых киназ и тирозинкиназ могут оказывать влияние на активность различных изоформ аденилатциклазы. В этом контексте мы считали целесообразным рассмотреть регуляцию различными киназами и протеинфосфатазами каждой изоформы аденилатциклазы по отдельности.
Данные, касающиеся влияния ПКС на активность АЦ I, достаточно противоречивы. Так, одни авторы наблюдали увеличение форсколин- и кальмодулин-стимулированной активности АЦ I в ответ на активацию протеинкиназы С [27], а другие нет [33]. Фосфорилирование Ser 545 и Ser 552 С1в домена АЦ I Са2+/кальмодулин-зависимой киназой IV влияет на Са2+/кальмодулин-индуцируемую активацию АЦ I без изменения базальной и форсколин-стимулируемой каталитической активности. Замещение любого из этих двух остатков серина на аланин предотвращает ингибирование АЦ I Ca2+/кальмодулин-зависимой киназой IV. Эти данные предполагают, что Ca2+/кальмодулин-зависимая киназа IV может функционировать как регулятор отрицательной обратной связи для АЦ I, регулируя уровень цАМФ в клетках [25].
Обработка клеток НЕК 293, экспрессирующих АЦ II, форболовыми эфирами приводит к активации фермента почти в 10 раз. Стауроспорин (ингибитор ПКС) предотвращал действие форболовых эфиров на синтез цАМФ АЦ II. Эти данные свидетельствуют в пользу того, что эта изоформа аденилатциклазы регулируется ПКС [33]. Обработка АЦ II ПКСa приводит к повышению базальной, форсколин - стимулируемой и bg-стимулируемой активностей, а также значительно повышает чувствительность к стимуляции a-субъединицей Gs белка [35]. Анализ in vivo, использующий химерные аденилатциклазы I/II, а также мутационный анализ позволили определить область в С2a АЦ II (1034-1068), которая содержит Thr 1057, фосфорилируемый ПКС [17]. Вместе с тем, следует отметить, что Taussig и Gilman [28] не сумели обнаружить фосфорилирования АЦ II, экспрессируемой в клетках Sf 9, при добавлении активированной ПКС.
АЦ III и Са2+/кальмодулин-зависимая протеинкиназа II являются коэкспрессированными в цилиарных ресничках ольфакторных нейронов. Поэтому кальмодулин-зависимая протеинкиназа II потенциально может регулировать активность АЦ III в процессе ольфакторной сигнализации. Действительно, было показано, что фосфорилирование Са2+/кальмодилин-зависимой протеинкиназой II Ser 1076 приводит к ингибированию АЦ III; мутация Ser 1076 на аланин превращает АЦ III в полностью нечувствительную к ингибированию протеинкиназой II. Фосфорилирование АЦ III по Ser 1076 Са2+/кальмодулин-зависимой киназой II значительно усиливается в нейронах после стимуляции одорантами. Ингибиторы Са2+/кальмодулин-зависимой протеинкиназы II, блокируя фосфорилирование АЦ III, предотвращают снижение повышенного уровня цАМФ, вызванного одорантами. Эти данные устанавливают новый механизм для окончания альфакторной сигнализации, который может иметь важное значение в ответах на одоранты [21].
Обработка АЦ IV ПКСa оказывала незначительное увеличение базальной и форсколин-стимулируемой активности, однако значительно снижала Gsa-cтимулируемую и bg-потенциирующую активность этой изоформы [35]. Эти данные указывают на то, что протеинкиназы могут изменять как активность различных изоформ аденилатциклазы, так и их чувствительность к регуляции различными субъединицами G белка.
Kawabe и соавт. [11] фосфорилировали АЦ V in vitro, используя ПКС, и продемонстрировали заметное увеличение ферментативной активности. Эффект был специфичен для a и s изоформ ПКС, предполагая перекрестную связь между этой изоформой аденилатциклазы и Gq - опосредованным (ПКСa), а также тирозинкиназными (ПКСs) сигнальными путями. Несмотря на достаточно сильный характер ответа, наблюдаемый in vitro, в условиях in vivo наблюдались достаточно умеренные эффекты активации АЦ V ПКС [27]. АЦ V фосфорилируется и ингибируется ПКА. Это ингибирование связано со снижением максимальной скорости фермента [24].
За исключением АЦ VI, остатки, фосфорилируемые в аденилатциклазах различными киназами, локализованы только в одном цитозольном домене (либо в С1в, либо в С2) [25]. Фосфорилирование множественных остатков АЦ VI ПКС уменьшает форсколин-индуцированную ферментативную активность [29,22]. Регуляции АЦ VI фосфорилированием представляет собой уникальный случай, так как эта изоформа может регулироваться тремя разными киназами, включая ПКА, ПКС и тирозинкиназу [1,29]. Особый интерес представляет тот факт, что ПКС и ПКА фосфорилируют один и тот же остаток Ser 674 в С1в домене АЦ VI [1,22]. Фосфорилирование АЦ VI ПКА уменьшает низкоаффинную стимуляцию Gsa субъединицей [1], в то время как фосфорилирование АЦ VI ПКС уменьшает ее каталитическую активность без изменения ее аффинности к форсколину и Gsa [29]. Это различие во влиянии ПКС- и ПКА-опосредованного фосфорилирования на ферментативные свойства АЦ VI может быть обусловлено тем, что ПКС в дополнение к Ser 674, который является также сайтом фосфорилирования ПКА, фосфорилирует еще, по меньшей мере, три остатка (Ser 10, N-концевой домен; Ser 568, С1в домен и Thr 931, С2 домен) [29,22]. Хотя одиночные мутации Ser 10, Ser 568 и Ser 674 лишь частично ослабляют ПКС-опосредованное ингибирование АЦ VI, одновременная мутация двух или всех трех этих остатков резко увеличивает нечувствительность мутантных форм АЦ VI к обработке ПКС [22].
Другим остатком существенным для ПКС-опосредованного ингибирования АЦ VI является Thr 931 (С2 домен). В противоположность эффектам, которые наблюдались для одиночных мутантов АЦ VI (АЦ VI - S10A, АЦ VI-S568A и АЦ VI S674A), замещение Thr 931 аланином приводило к полной функциональной нечувствительности АЦ VI к ПКС-опосредованному ингибированию [22]. Основываясь на этих результатах Lin и соавт. [22] предполагают, что три цитозольных домена АЦ VI формируют сложный регуляторный комплекс. Фосфорилирование различных сайтов этого регуляторного комплекса может индуцировать тонкую настройку относительной ориентации каталитического ядра, приводящей к ингибированию ферментативной активности АЦ VI.
Интересно, что три другие изоформы аденилатциклазы (АЦ V, АЦ VIII и АЦ IX) содержат в позиции эквивалентной Thr 931 АЦ VI потенциальные сайты фосфорилирования ПКС [22]. Напомним, что АЦ V стимулируется ПКС [11]. Является ли сайт фосфорилирования АЦ V ПКС эквивалентным Thr 931 АЦ VI остается неизвестным. До сих пор неизвестны также протеинкиназы, оказывающие влияние на активность АЦ VIII и АЦ IX.
Поскольку каталитические ядра АЦ V и АЦ VI высокогомологичны между собой, значительный интерес представляет тот факт, что ПКС оказывает противоположные функциональные эффекты на эти изоформы. Как указывалось выше, по меньшей мере две различные изоформы ПКС могут фосфорилировать АЦ V и усиливать ее каталитическую активность [11]. Среди сайтов фосфорилирования АЦ V ПКС пять находятся в позициях, эквивалентных таковым в АЦ VI (Ser 145, Ser 325, Ser 489, Ser 568 и Thr 931).
Два же основных сайта фосфорилирования ПКС в АЦ VI (Ser 10 и Ser 674) отсутствуют в АЦ V. Важность Ser 674 является очевидной, поскольку этот остаток может фосфорилироваться как ПКС, так и ПКА в АЦ VI [1,22], а в АЦ V он может фосфорилироваться только ПКА, вызывая ингибирование фермента [24]. Возникновение отрицательных зарядов, обусловленное фосфорилированием Ser 674 в АЦ VI и эквивалентного сайта в АЦ V, может угнетать каталитическую активность этих изоформ. Отсутствие ПКС-опосредованного фосфорилирования Ser 674 в АЦ V, по-видимому, способствует отсутствию негативного эффекта ПКС на АЦ V. Кроме того, в N-концевом домене АЦ VI ПКС фосфорилирует Ser 10, а в АЦ V такой сайт фосфорилирования ПКС отсутствует.
Имеются единичные сведения о том, что обработка АЦ VII протеинкиназой С сопровождается увеличением циклазной активности[15].
По-видимому, АЦ VIII является единственной изоформой мембраносвязанных аденилатциклаз млекопитающих, активность которой не зависит от процессов фосфорилирования/дефосфорилирования. Так, ее активность не изменяется под влиянием ПКС [14] и Са2+/кальмодулин-зависимых протеинкиназ II и IV [25]. Данных о влиянии ПКА на активность АЦ VIII нами не обнаружено.
АЦ IX ингибируется ионами Са2+ и этот эффект селективно блокируется циклоспорином А и FR 506, то есть специфическими блокаторами Са2+/кальмодулин-зависимой протеинфосфатазы 2В - кальцинейрина. Кальцинейрин, являясь сенсором внутриклеточного кальция, опосредует отрицательную обратную связь влияния внутриклеточного кальция на рецептор-стимулируемое образование цАМФ в различных клетках [9]. Значительная экспрессия АЦ IX в гиппокампе и жизненно важная роль регуляции цАМФ кальциевым сигналом в синаптической пластичности свидетельствуют о том, что АЦ IX может быть важной физиологической мишенью для протеинкиназного и протеинфосфатазного каскадов, которые поддерживают долговременную модуляцию синаптической передачи [3].
Регуляция активности мембраносвязанных аденилатциклаз млекопитающих на уровне мембранных доменов М1 и М2
Обнаружение того факта, что цитоплазматические домены С1 и С2 аденилатциклаз, экспрессируемые в бактериях, могут образовывать активную каталитическую форму фермента in vitro, которая подвержена регуляторному влиянию многих агентов, образно говоря, отодвинуло исследование роли мембранных доменов на второй план. Тем не менее, можно выдвинуть ряд соображений относительно того, что функция мембранных доменов М1 и М2 не ограничивается только заякориванием аденилатциклазы в мембране. Во-первых, размеры мембранных доменов, составляющие около 30% от величины всего фермента, намного превышают необходимые для заякоривания аденилатциклазы в мембране. Иными словами, для заякоривания фермента в мембране достаточно одного/двух трансмембранных сегментов, а не двенадцати. Во-вторых, домены М1 и М2 сильно отличаются друг от друга по аминокислотной последовательности и все попытки сравнить между собой эти домены окончились неудачей из-за их низкой консервативности. Так, сравнительный анализ аминокислотных последовательностей М1 и М2 АЦ V кролика показывает только 12% их идентичности [26]. Кроме того, домены М2 на 54-87 аминокислот длиннее, чем изоформа-соответствующие домены М1. Дополнительные аминокислотные остатки доменов М2 участвуют в формировании более длинных последовательностей внутри- и экстраклеточных петель с неизвестной функцией. В третьих, независимо от классификации девяти мембраносвязанных аденилатциклаз млекопитающие на четыре сходно регулируемые группы (типы I, III, VIII; II, IV, VII; V, VI; IX) их мембранные домены указывают на почти полное отсутствие консервативности. Это ставит вопрос о том, обусловлены ли эти различия последовательностей мембранных доменов отсутствием эволюционного давления на сохранение функциональной специфичности или непознанными до сих пор функциональными требованиями. В четвертых, сравнительный анализ аденилатциклаз любой индивидуальной изоформы, полученных от животных, находящихся на различных уровнях эволюционного развития, указывает на высокую степень гомологии соответствующих мембранных доменов. Например, при сравнении АЦ V кролика и крысы наблюдается идентичность их последовательностей на 99% в областях цитоплазматических доменов С1 и С2 и на 92% в мембранных якорях. Сравнительный анализ аминокислотных последовательностей АЦ V кролика и АЦ VII крупного рогатого скота указывает на снижение идентичности до 53% для каталитических доменов и до 24% для соответствующих мембранных якорей [26]. Это является доказательством отчетливых изофермент-специфических функций доменов М1 и М2. В пятых, структурные ограничения, проявляемые доменами М1 и М2, по-видимому, влияют на взаимодействие между цитоплазматическими доменами С1 и С2, поскольку мутации внутри мембранных якорей приводят к уменьшению или вообще к полному отсутствию аденилатциклазной активности [17].
Первые биохимические свидетельства возможной изоформа-специфической функции аденилатциклазных мембранных доменов в образовании каталитически активного псевдогетеродимера были представлены в 2001 году [26].
Авторы использовали каталитические домены (С1 и С2) из АЦ V и мембранные домены (М1 и М2) из аденилатциклаз V и VII типов для конструирования ферментов с дуплицированными, инвертированными, полностью замененными и химерными мембранными якорями. Все разновидности этих ферментов были экспрессированы в клетках НЕК 293. Полученные данные показали, что ферментативная активность была обнаружена только в том случае, когда оба мембранных домена М1 и М2 были получены из какой либо одной изоформы, то есть либо из АЦ V, либо из АЦ VII. Более того, мембранные домены М1 и М2 из АЦ VII были столь же эффективны для сохранения аденилатциклазной активности каталитических доменов АЦ V, как и мембранные домены М1 и М2 из АЦ V. Конструкции аденилатциклаз с химерными мембранными доменами, то есть М1 из АЦ V, а М2 из АЦ VII (или, наоборот) были существенно инактивированными, и не активировались форсколином и Gsa, хотя уровни экспрессии и мембранная локализация в клетках этих ферментов была нормальной. Эти данные указывают на функционально важную роль мембранных доменов аденилатциклаз. Мембранные домены М1 и М2, по-видимому, взаимодействуют, спариваясь специфически, только в пределах данной изоформы аденилатциклазы, то есть изоформа-специфическим способом. Это взаимодействие непосредственно связано с образованием каталитической поверхности раздела между цитоплазматическими доменами С1 и С2 аденилатциклазы. Предполагается, что белок-белковые взаимодействия аденилатциклазных мембранных доменов могут составлять новый, все еще не изученный уровень регуляции аденилатциклазы.
На наш взгляд, еще одним механизмом регуляции аденилатциклазной активности, осуществляемым через влияние на М1/М2 взаимодействие, является гликозилирование экстраклеточных петель аденилатциклаз. У всех мембраносвязанных аденилатциклаз млекопитающих потенциальные сайты N-гликозилирования локализованы в области экстраклеточных петель 5 и/или 6 [20]. Действительно, некоторые изоформы аденилатциклазы (АЦ II, АЦ III, АЦ VI, АЦ VIII) являются гликозилированными [20,5]. Гликозилирование АЦ VIII влияет на каталитическую активность без изменения значения Кm и ее чувствительности к кальмодулину [5]. Интересно отметить, что только полностью гликозилированная форма АЦ III (в отличие от негликозилированной формы) может быть фосфорилирована Са2+/кальмодулин-зависимой киназой II [32]. Wu и соавторы [20] показали, что АЦ VI является гликозилированной in vivo. Обработка клеток НЕК 293, экспрессирующих АЦ VI, туникамицином, который блокирует N-гликозилирование, приводит к значительному снижению стимуляции АЦ VI форсколином, но не Gsa по сравнению с гликозилированной формой фермента. Кроме того, было показано, что гликозилирование АЦ VI не только влияет на ее каталитическую активность активатор-зависимым способом, но также изменяет способность к регуляции протеинкиназой С и рецептором, связанным с Gi белком [20].
|
|
|
|
Дата добавления: 2014-01-11; Просмотров: 1462; Нарушение авторских прав?; Мы поможем в написании вашей работы!