КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Коферменти, що використовуються при диханні
| Кофермент | Функції | Вітамін, з якого синтезується кофермент |
| НАД+ (NAD+) – нікотинамідаденіндинуклеотид | Передає атоми водню в ланцюг переносу електронів | В3 (ніацин, нікотинова кислота) |
| ФАД (FAD) – флавінаденіндинуклеотид | Передає атоми водню в ланцюг переносу електронів | В2 (рибофлавін) |
| КоА (СоА) – кофермент А | Переносить ацетильну групу в ЦТК | В5 (пантатенова кислота) |
4.2 Обмін речовинами між клітиною й навколишнім середовищем
Клітина, будучи відкритою системою, обмінюється речовинами з навколишнім середовищем. Пасивний перенос відбувається за рахунок кінетичної енергії часток, що пересуваються, тоді як для активного транспорту необхідна метаболічна енергія.
Плазматична мембрана сповільнює й регулює обмін речовинами. Деякі з них вона просто пропускає (виборча проникність), а інші активно перекачує навіть проти градієнта концентрації.
При вільному транспорті молекули або іони пересуваються (пасивно) у вільному виді, при транспорті з переносником - у комплексі з наявними в самій мембрані транспортними молекулами, які називають переносниками (пасивно або активно). При ендо- і екзоцитозі активно транспортуються частки або крапельки рідини, видимі в мікроскоп.
4.2.1 Вільний транспорт
У газах і рідинах молекули й іони перебувають у постійному безладному тепловому русі. Якщо в газовій суміші або розчині існує градієнт концентрації якоїсь речовини, то внаслідок теплового руху концентрація його поступово вирівнюється. При цьому рух стає статистично спрямованим, і його називають дифузією. Розчинена речовина переміщається по напрямку власного градієнта, оскільки там, де більше розчинених молекул, число молекул води в одиниці об'єму ("концентрація води") зменшено. Дифузія - вирівнюючий процес, що збільшує ентропію, тому що веде до менш упорядкованого й тому більше ймовірному стану.
Дифузія через мембрану відбувається більш повільно, тому що ліпіди мембрани служать перешкодою, яке обмежує швидкість. Речовини, розчинні в ліпідах, можуть дифундувати через ліпідний шар, у той час як іншим речовинам доводиться використати малюсінькі "пори" у шарі ліпідів.
Дифузія, зокрема дифузія через мембрану, відбувається в напрямку хімічного потенціалу, що залежить від температури й концентрації. На заряджені частки (іони), крім того, впливають електричні потенціали (наприклад, мембранний потенціал). Їхня дифузія йде по напрямку електрохімічного потенціалу. Вода переміщається по градієнту водного потенціалу.
Осмос - це дифузія води через напівпроникну мембрану, наприклад плазматичну. Такі мембрани добре пропускають воду, але малопроникні для розчинених у ній речовин. Якщо помістити клітину в чисту воду, то створюється градієнт водного потенціалу: зовні 100% води, а усередині менше 100% (інше - розчинена речовина). Тому вода переходить усередину по градієнту своєї концентрації (точніше, водного потенціалу). Вода зв'язується капілярними силами або ж у вигляді гідратаційної води. Осмотичне поглинання води веде до збільшення об’єму клітини.
У зрілих рослинних клітинах головним "осмотичним простором" є центральна вакуоль. Завдяки міцній й еластичній стінці клітина лише ненабагато збільшується в розмірах, і осмотичне поглинання води веде до створення високого гідростатичного тиску у вакуолі - тургорного тиску.
4.2.2 Транспорт із переносником
Плазматична мембрана містить транспортні білки, які як переносники зв'язують субстрати й транспортують їх через мембрану. Протомери цих олігомерних білків утворюють гідрофільний канал, що закривається (пору з "клапаном", рис. 4.3). При зміні конформації ділянка, що зв'язує субстрат, переміщається з однієї сторони мембрани на іншу й у такий спосіб переносить субстрат. Існують різні переносники з різним механізмом дії й різною субстратною специфічністю.
| А | 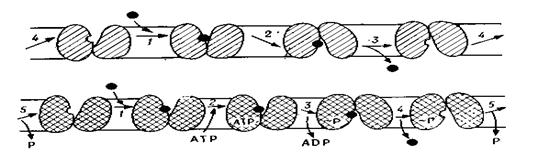
|
| Б |
Рис. 4.3. Транспорт із переносником. A. Каталізуєме проходження через мембрану, модель. Б. Активний транспорт, модель.
Каталізуємий переніс (полегшена дифузія) - це пасивний транспорт із переносником по градієнту електрохімічного потенціалу. Зв'язування субстрату веде до зміни конформації переносника. Таким чином, цукри, амінокислоти й інші речовини проходять через мембрану, звичайно мало проникну для них. Це головний механізм виборчої проникності мембран.
Особливий випадок каталізованого переносу - сполучений транспорт. Деякі переносники транспортують два різні субстрати разом або в одному напрямку (паралельний транспорт, наприклад перенос протонів і лактози в Escherіchіa colі, або в протилежних напрямках (антипаралельний транспорт), зокрема перенос Н+ й Na+ через мембрани мітохондрій. Активний транспорт здійснюється проти градієнта хімічного або електрохімічного потенціалу. Розщеплення АТР веде до зміни конформації транспортної АТР ази, що часто буває пов'язане з її тимчасовим фосфорилюванням. Цей процес служить "двигуном" активного транспорту. Таким способом через мембрану переносяться протони (протонний насос) або неорганічні іони (іонний насос).
4.3 Дисиміляція як джерело енергії
Дисиміляція являє собою розщеплення органічних субстратів з використанням їхньої хімічної енергії.
4.3.1 Огляд процесів дисиміляції
При диханні субстрат без залишку розщеплюється до бідних енергією неорганічних речовин з відповідно високим виходом енергії. При шумуванні субстрат руйнується не повністю - до органічного кінцевого продукту, ще багатого енергією, і вихід енергії тут відповідно невеликий. Дихання - аеробний окисний процес, для нього необхідний кисень. Шумування - анаеробні процеси. Вищі тварини й рослини дихають. Шумування властиво головним чином мікроорганізмам (бактеріям, дріжджам), але при недоліку кисню може зустрічатися також у клітинах вищих рослин і тварин.
Найважливішими субстратами для дихання й більшості шумувань служать вуглеводи. Крім того, при диханні можуть використовуватися жири й білки. Субстратами для шумування можуть також бути спирти, органічні кислоти й інші речовини. Через малий вихід енергії клітини, що здійснюють шумування, повинні витрачати більші кількості субстрату, чим дихаючі клітини.
Дихання складається із двох часткових процесів:
1) поступового розщеплення субстрату з відібранням водню, що зв'язується з коферментами (символ [Н2]), і
2) поступового окислювання [Н2] у результаті переносу його на кисень.
Для вуглеводів:
| Розщеплення субстрату: C6H12О6 + 6Н2О ® 6CО2+ 12[Н2] (4.3) Окислення водню: 12[H2]+ 6О2 ®12Н2О (4.4) |
| С6Н12О6 + 6Н2О + 6О2 ®6СО2 + 12Н2О (4.5) DG0' = –2875 кДж/моль |
Молочнокисле шумування(м'язи, бактерії Lactobacillus, Streptococcus):
С6Н12О6 ® 2СН3СН-ОН-СООН; DG0' = – 200 кДж/моль (4.6)
Спиртове шумування(Saccharomyces та інші дріжджі):
С6Н12О6 ® 2СН2СН2ОН + 2СО2 ; DG0' = –235 кДж/моль (4.7)
Маслянокисле шумування (Clostridium та інші бактерії):
С6Н12О6 ® С3Н7СООН + 2СО2 + 2Н2; DG0' = –265 кДж/моль (4.8)
В інших бактерій зустрічається мурашинокисле й пропіоновокисле шумування.
4.3.2 Шляхи розщеплення вуглеводів
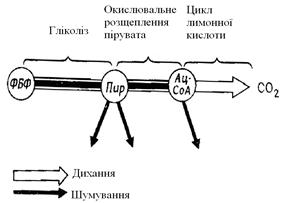
При розщепленні субстрату на нього послідовно впливають різні ферменти. Деякі етапи цього процесу при диханні й шумуванні ідентичні. Розщеплення вуглеводів починається із гліколізу, після чого шляхи дихання й шумування розходяться.
Гліколіз - це процес окисного розщеплення, що відбувається в основній речовині цитоплазми (і клітинного ядра) і ведучий від фруктозо-1,6-біфосфата до пірувату (піровиноградної кислоти СН3СО-СООН) - проміжного продукту, що має велике значення (рис. 4.4).
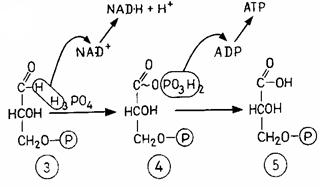
Фруктозо-1,6-біфосфат спочатку розщеплюється на два ізомерних продукти, що перебувають у рівновазі між собою. Один з них, 3-фосфогліцеринальдегід, у результаті одного етапу окислювання перетворюється у фосфогліцерат, з якого потім утвориться піруват. Окислювання каталізує дегідрогеназа (фермент, що відщеплює водень).
| 
|
| А | В |

| |
| Б |
Рис. 4.4. Гліколіз. А. Місце в обміні речовин при диханні й шумуванні. ФБФ - фруктозо-1,6-біфосфат; Пір - піруват; Ац-Соа - ацетил-Соа.
Б. Хімічні етапи. В. Окисна частина ланцюга гліколізу з фосфорилюванням на рівні субстрату. 1 - фруктозо-1,6-біфосфат; 2 - дигідроксиацетонфосфат; 3 - 3-фосфогліцеринальдегід; 4 - 1,3-дифосфогліцерат; 5 - 3-фосфогліцерат; 6 - 2-фосфогліцерат; 7 - фосфоенолпіруват; 8 - піруват (енольна форма й кето-форма)
Водень приєднується до коферменту NAD+ (нікотинамідаденіндинуклеотид, вітамін В3) (рис. 4.5). Відновлений кофермент NAD-H - один з найбільш універсальних переносників водню. Щоб енергію окиснювання можна було зберегти у формі АТР, вона використається спочатку для "фосфорилювання на рівні субстрату"; при цьому утворюється високоенергетичний проміжний продукт 1,3-дифосфогліцерат. Потім фосфатна група (разом з енергією) переноситься на ADP, і в результаті останній перетворюється у високоенергетичну сполуку АТР. Продуктами гліколізу є піруват, водень у формі NAD-H й енергія у формі АТР.
При різних видах шумування подальша доля продуктів гліколізу - пірувату й NAD-H - різна. При молочнокислому шумуванні водень переноситься на піруват й у результаті відразу утворюється лактат (молочна кислота):
СН3СО-СООН + NAD-H + Н+ ® СН3СН-ОН-СООН + NAD+ (4.9)
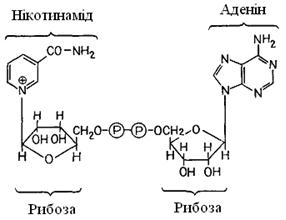
Рис. 4.5. NAD +, структурна формула
При спиртовому шумуванні піруват спочатку декарбоксилюється, тобто від нього відщеплюється СО2, а потім проміжний продукт ацетальдегід відновлюється в етанол у результаті переносу водню:
СН3СО-СООН ® СО2 + СН3СНО (4.10)
СН3СНО + NAD-H + H+ ® СН3СН2ОН + NAD+ (4.11)
Функція NAD + і подібних з ним переносників водню полягає в тому, щоб у першій реакції приймати водень (відновлюватися), а в другій реакції віддавати його (окиснюваись).
При диханні водень, одержаний в результаті гліколізу, транспортується до кисню. Піруват піддається окисному декарбоксилюванню при участі мультиферментного комплексу (комплекс білків, що здійснюють кілька ферментативних функцій) і декількох коферментів. Один кофермент приймає відщиплений водень (окислювання), а інший - кофермент A (CoA-SH, вітамін В5) - приєднує ацетильну групу, що залишилася (-CO-CH3). CoA-SH містить реакційно-здатні сульфгідрильні групи (–SH) (рис. 4.6).
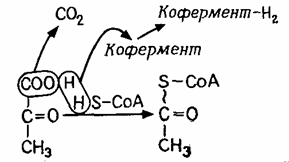 |
Рис. 4.6. Розщеплення пірувату - окисне декарбоксилювання
Високоенергетична сполука ацетил- Cоа (CoA-S ~CO-CH3) ("активована оцтова кислота") є найважливішим проміжним продуктом клітинного метаболізму, що може мати різне походження й по-різному використовуватися.
При диханні ацетильний залишок повністю розщеплюється в циклі лимонної кислоти в результаті окислювання (відібрання Н), декарбоксилювання (відщіплення СО2) і гідратації (приєднання Н2О) (рис. 4.7). Загальне рівняння:
CoA-S ~CO-CH3 + 3Н2О ® CoA-SH + 2CО2+4[H2] (4.12)
Перед розщепленням ацетильний залишок зв'язується з оксалоацетатом; у результаті утворюється цитрат, що поступово розщеплюється, поки знову не залишається оксалоацетат; останній вступає в реакцію з новою молекулою ацетил- СоА, і цикл повторюється.
Одержуваний у цьому циклі водень приєднують коферменти, насамперед NAD.
Окислювання пірувату й цикл лимонної кислоти здійснюються в мітохондріях.
В основній речовині цитоплазми можливий ще інший шлях розщеплення вуглеводів, без гліколізу й циклу лимонної кислоти, - пентозофосфатний цикл. Глюкозофосфат піддається окисному декарбоксилюванню до пентозофосфата, а потім 6 молекул пентозофосфата перетворюються через ряд проміжних етапів в 5 молекул глюкозофосфату. Загальне рівняння (без фосфату):
6С6Н12О6 + 6Н2О ® 6С5Н10О6 + СО2 + 12 [H2] (4.13)
6С5Н10О5 ® 5С6Н12О6 (4.14)

Рис. 4.7. Цикл лимонної кислоти (цикл трикарбонових кислот - ЦТК). 1 - оксалоацетат; 1а - ацетил- СоА; 2 - цитрат; 3 - ізоцитрат; 4- оксалосукцинат; 5- -кетоглутарат; 6 - сукцинат (бурштинова кислота); 7 - фумарат; 8 - малат (яблучна кислота)
С6Н12О6 + 6Н2О ® 6СО2+ 12[Н2] (4.15)
Згідно хеміосмотичній гіпотезі, потік електронів насмоктує протони через мітохондріальну мембрану, і електрохімічний протонний потенціал, що при цьому створюється, доставляє енергію для утворення АТР. Відповідно до цієї гіпотези, у дихальному ланцюзі чергуються оксидоредуктази, що переносять електрони (е-), або електрони ы водень (е -+Н+). Вони розташовані в мембрані так, що прийняття Н+ може відбуватися тільки на внутрішній, а віддача - тільки на зовнішній стороні.
4.3.3 Біологічне окиснювання
Основне джерело енергії для синтезу АТР при диханні - це окислювання киснем водню (пов'язаного з коферментом), одержуваного в результаті гліколізу, окислювання пірувату й реакцій циклу лимонної кислоти, а іноді й пентозофосфатного циклу.
Кожна реакція окислювання (віддача електронів) сполучена з відновленням (прийняттям електронів), так що ми завжди маємо справу з окислюванням-відновленням.
Н2 ® 2Н+ + 2 е – (окислювання)
0,5×О2 + 2 е – ® О2– (відновлення)
H2+O2 ® 2Н++ О2- = Н2О (окислювання-відновлення)
Системи Н2/2Н+, О2–/0,5·О2 є окислювально-відновними системами. Окислювання-відновлення - це перенос електронів від однієї окислювально-відновної системи до іншої. Окислювально-відновні системи з більшою спорідненістю до електронів ("акцептори електронів") мають схильність приймати електрони (наприклад, О2-® 0,5×О2+2 е -), а з малою спорідненістю ("донори електронів") - віддавати їх (Н2 ® 2Н++ 2 е -). Перехід електронів від речовини з більш низькою спорідненістю до речовини з більш високою спорідненістю до електронів (від донора до акцептора) - екзергонічний процес, що протікає мимовільно з виділенням енергії. Перенос електронів у протилежному напрямку - ендергонічний процес.
Спорідненість до електронів виміряється як окислювально-відновний потенціал (ОВП). Негативний ОВП означає низьку спорідненість до електронів, позитивний - високу спорідненість.
Для системи Н2/2Н+ Е'0 = - 0,42 В, для системи О2-/0,5·О2 Е'0 = +0,81В. Мимовільний, екзергонічний перенос електронів відбувається по напрямку від більш негативного до більш позитивного окислювально-відновного потенціалу:
Н2/2Н+ ® О2-/0,5·О2
Чим більше різниця в ОВП між двома окислювально-відновними системами, тим більша кількість енергії (–DG0') звільняється.
Окиснювально-відновна система NAD-H/NAD має ОВП - 0,32 В. Різниця потенціалів стосовно кисню (Е0' = 0,81 В) велика: DЕ'0 = 1,13 В, звідки DG°' = 218 кДж/моль. Між NAD-H і О2 розташовується ланцюг транспорту електронів, або ланцюг дихання. Такі ланцюги складаються з ряду окислювально-відновних систем, які послідовно передають один одному електрони. Завдяки цьому велика різниця ОВП (1,13 В) дробиться, так само як і вивільнювана енергія, на декілька "роздавальних пунктів", де енергія може передаватися системі ADP-АТР.
Дихальний ланцюг складається з оксидоредуктаз (рис. 4.8) - дегідрогеназ, флавопротеїдів, убіхінона, цитохромів і білків, що містять залізо й сірку. Деякі із цих білків транспортують тільки електрони (е-), інші - водень (е- +Н+).
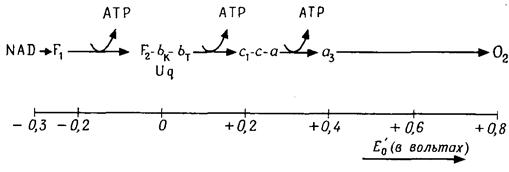
Рис. 4.8. Ланцюг дихання. Розташування й окислювально-відновні потенціали компонентів ланцюга дихання (спрощене). F1, F2 – флавопротеїди FpDl и FpD2; Uq – убіхінон; от bK до а3 – цитохроми
В основному вони утворюють мультиферментні комплекси, будучи інтегральними білками внутрішньої мембрани мітохондрій; тільки більша частина дегідрогеназ слабко пов'язана із внутрішньою стороною мембрани: це її периферичні білки. Дегідрогенази переносять водень із субстрату на свій кофермент, у більшості випадків NAD+, що слабко пов'язаний з ферментом у якості його косубстрата:
Субстрат – Н2 + NAD+ ® Субстрат + NAD-H + H+ (4.16)
Флавопротеїди в якості простетичної групи використовують FAD, рідше FMN (флавінмононуклеотид). Обидва ферменти містять рибофлавін (вітамін В2):
NAD-H + H+ + FAD ® NAD+ + FAD - Н2 (4.17)
Водень від NAD-H послідовно переходить до двох флавопротеїдів - FpDl й Fp2 (рис. 4.8). Інші флавопротеїди є дегідрогеназами й приймають водень безпосередньо від субстрату (наприклад, від сукцината):
Субстрат-Н2+ FAD ®Субстрат + FAD-Н2 (4.18)
Убіхінон(Uq, кофермент Q) Передбачувана реакція:
FAD-Н2 + Uq ® FAD + UqH2 (4.19)
Цитохроми використовують як коферменти залізопорфірини (геми), що переносять електрони. Порфірини являють собою кільцеві структури із чотирьох пірольних кілець із центральним атомом металу (рис. 4.9). У цитохромі центральний атом заліза шляхом зміни своєї валентності здійснює перенос електронів: Fe3+ + e – ® Fe2+.
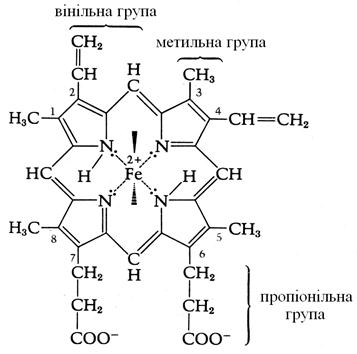
Рис. 4.9. Простетична група цитохромів
Дихальний ланцюг містить у тварин цитохроми bк, bт, с1, с, а і а3. У рослин відмінності невеликі, у бактерій більш значні. При переході електронів від цитохрому до цитохрому валентність заліза увесь час змінюється.
Білки, що містять залізо й сірку, у яких залізо зв'язане через S, перебувають у різних ділянках ланцюга дихання разом із флавопротеїдами й цитохромами і беруть участь у транспорті електронів зі зміною валентності Fe.
Енергія, що звільняється при екзергонічній дисиміляції, зберігається (незважаючи на великі втрати у вигляді тепла) у формі енергії АТР у результаті ендергонічного фосфорилювання ADP. Фосфорилювання на рівні субстрату відбувається при окислюванні різних субстратів дегідрогеназами. Фосфорилювання в дихальному ланцюзі відбувається при окислюванні одного компонента дихального ланцюга наступним компонентом. При анаеробному шумуванні фосфорилювання на рівні субстрату - єдине джерело енергії для синтезу АТР.
У ланцюзі дихання при русі протона від NAD-Н до O2 звільняється 218 кДж на 1 моль NAD-H. Із цієї кількості шляхом утворення 3 молів АТР запасається при стандартних умовах 90 кДж, а в клітині, імовірно, 120 кДж. Ферментний комплекс, що утворює АТР, - мембранна АТР аза, перебуває на внутрішній стороні внутрішньої мітохондріальної мембрани.
Процес клітинного дихання можна розділити на три послідовних реакції: гліколіз, цикл лимонної кислоти (ЦТК), ланцюг переносу електронів (ЦПЕ) (рис. 4.10). Кількість АТР, синтезованої при розщепленні глюкози при диханні становить 38 молекул на кожну молекулу глюкози (табл. 4.3).
4.4 Асиміляція
Асиміляція - це перетворення чужорідних речовин у компоненти власного організму. Автотрофна асиміляція буває двох видів - фотосинтез і хемосинтез (табл. 4.4). Зелені рослини, синьо-зелені водорості й деякі бактерії, що синтезують органічні речовини з неорганічних - "первинні продуценти", мають величезне значення для життєдіяльності всіх живих істот. Гетеротрофна асиміляція інших організмів - порівняно більш простий процес перетворення одних органічних речовин в інші.
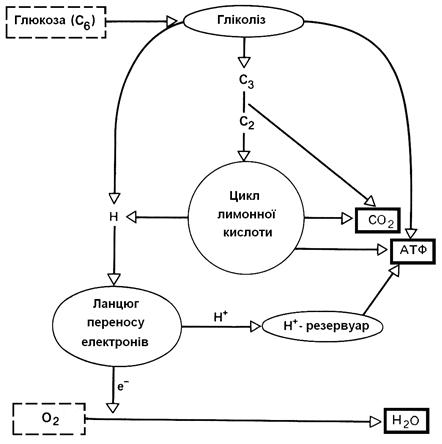
Рис. 4.10. Розщеплення глюкози в процесі дихання
Тому що органічні речовини являють собою сполуки вуглецю, то вирішальне значення має асиміляція вуглецю. Це процес відновлення, що веде від максимально окисленої вихідної речовини СО2
Таблиця 4.3
|
|
Дата добавления: 2014-01-11; Просмотров: 1164; Нарушение авторских прав?; Мы поможем в написании вашей работы!