КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Экология сообществ и экосистем 3 страница
|
|
|
|
Еще более сложные коадаптации между растениями и животными формируются при эндозоохории. У большинства птиц и у многих млекопитающих семена, пройдя через пищеварительный тракт, сохраняют жизнеспособность и в результате транспортируются на довольно далекие расстояния. В Норвегии подсчитали, что со свежим конским навозом, вносимым на поля как удобрение из расчета 60 т/га, засевается на каждом гектаре до 900000 сорных растений. Так распространяются семена ярутки (Thlaspi arvense). Овцы, лошади и другие домашние животные разносят пастушью сумку (Capsella bursa-pastoris), овсюг (Avena fatua), горчицу (Sinapis), дикую редьку (Raphanus raphanistrum), мокрицу (Stellaria media). Воробьи разносят семена земляники, бузины, калины, крушины, жимолости и других ягодников. Большую роль в распространении растений играют перелетные птицы. Благодаря им растения северных областей попадают в степи. Это шиповник, черемуха, крушина и др. Дрозд деряба (Turdus viscivorus) переносит семена омелы. На Гавайских островах широко расселился сорняк лантана (Lantana camara) в связи с акклиматизацией там птицы майны (Acridotheres tristis). С деятельностью плодоядных птиц связывают появление большинства деревьев на океанических островах.
Иногда семена распространяются через несколько звеньев. К примеру, через рыбоядных птиц расселяются водные растения.
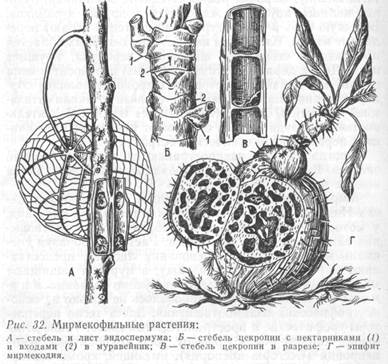
Мирмекофилия. «Грибные сады». В тропических лесах Индии, Китая и других стран встречаются растения, у которых исключительно своеобразные взаимоотношения складываются с муравьями. Растения, образуя специальные нектарники у основания листьев, предоставляют муравьям убежище и пищу, а муравьи защищают их от вредителей. Это явление получило название мирмекофилии. Оно сформировалось не только на основе трофических взаимоотношений. Здесь тесно переплетены трофические и пространственные связи. Такие отношения характерны, к примеру, для южноамериканской цекропии (Cecropia adenopus), имеющей, кроме нектарников, своеобразные полости внутри стебля, которые и заселяются муравьями. У эпифита мирмекодии (Myrmecodia pentasperma) вздутый стебель (ложная луковица) также пронизан полостями и представляет собой искусственный муравейник. У эпифита дисхидии (Dischidia rafflesiana) образуются листовые кувшинчики, где собирается влага, используемая самим растением. Некоторые из этих кувшинчиков оказываются перевернутыми вверх дном, и тогда их заселяют муравьи. У мексиканского кустарника Acacia cornigera муравьи поселяются в расположенных у основания листьев крупных полых колючках. Кроме того, на конце каждого листочка размещается особое тельце белого цвета, наполненное жиром и белком – прекрасным кормом для муравьев. Мирмекофильных растений в тропиках известно до 3000 (рис. 32).
|
|
|
К мирмекофилам относятся и многие животные, находящие в муравейниках укрытие и питающиеся отбросами муравьев, гниющим строительным материалом гнезда или превращающиеся в настоящих с и м φ и л о в, т. е. нахлебников или даже своеобразных паразитов и хищников муравьев. Известно свыше 2000 видов мирмекофильных членистоногих.
Между животными-мирмекофилами и муравьями формируются исключительно сложные взаимоотношения.
Симфилов муравьи кормят жидкой пищей, как и своих личинок. Симфилы же, благодаря наличию специфичных желез, выделяют вещества, которыми питаются муравьи. Мирмекофильными насекомыми являются, например, тропические пауссиды (Paussidae), жуки-ощупники (Pselaphidae), стафилиниды (Staphylinidae). Интересны взаимоотношения с муравьями у некоторых видов клещей семейства Antennophoridae. Они держатся на нижней стороне головы муравьев и слизывают с их щупиков и челюстей капельки пищи. Классическим примером мирмекофила является жук ломехуза (Lomechusa strumosa), встречающийся только в муравейниках. Иногда муравьи уделяют им так много внимания, что от этого страдают сами – семья их деградирует и нередко погибает. Сущность подобных взаимоотношений до конца еще не выяснена.
|
|
|
Группа беспозвоночных животных, ведущих аналогичный образ жизни в термитниках, получила название термитофилов.
Хотя биология мирмекофилов и термитофилов изучена слабо, все же форму связи их с муравьями можно охарактеризовать как своеобразный симбиоз, а в отдельных случаях как паразитизм.
Чрезвычайно сложные отношения сформировались у муравьев с некоторыми грибами. Обитающие в Центральной и Южной Америке муравьи-листорезы (Atta, Acromyjmex) срезают челюстями кусочки листьев, но не питаются ими и не строят из них жилье, а доставляют их в муравейник, тщательно пережевывают, смешивают с экскрементами и слюной и складывают в специальные камеры. На таких своеобразных грядках вырастают гифы грибов. На свободных концах гиф образуются многочисленные скопления коротких и вздутых ветвей, называемых кольраби. Они и служат пищей муравьям. Молодые самки, покидая муравейник, забирают с собой кусочки грибницы и при постройке нового муравейника сразу же приступают к возделыванию «грибных садов».
Известно до 100 видов таких муравьев. Очень мелкие Cyphomyrmex rhimosus поселяются в пустых раковинах или во вздутых листьях растений, а грибную плантацию размещают глубоко под землей, где выращивают желтые дрожжи, удобряя грядки экскрементами гусениц. Другие виды цифомирмексов возделывают грибные сады на компосте из древесной пыли и экскрементов жуков-дровосеков. Экскрементами гусениц удобряют свои плантации также муравьи Trachimyrmex и Apterostigma.
Установлено, что в гнездах муравьев произрастают такие грибы, как Hypomyces и шляпочный гриб Rhozites gongylophora, которые больше нигде не обнаружены. Только в муравейниках некоторых муравьев рода Lasius найден гриб Septosporium; гриб Leicosporium известен лишь из гнезд муравья Cyphomyrraex costatus. Все это позволяет сделать предположение, что в грибных садах муравейников в процессе естественного отбора образовались особые виды «муравьиных грибов».
Своеобразные грибные плантации встречаются также в гнездах термитов, где на специально откладываемых скоплениях экскрементов и кусочков древесины разводятся различные плесневые грибы. Но иногда, как и у муравьев, в них встречаются специфические грибы, которые больше нигде не произрастают (например, Termito-myces).
|
|
|
Грибные сады встречаются и у некоторых жесткокрылых. Так, самка непарного короеда (Xyleborus dispar), покидая гнездо, в специальных «карманчиках» уносит кусочки грибницы. Прогрызая ходы в новом дереве, короед образует там грибные камеры и рассеивает в них грибницу. Жук поддерживает в этих камерах определенную влажность – смачивает грибные грядки особыми выделениями, которые стимулируют рост полезных грибов и уничтожают сорняки. Личинки короеда питаются не древесиной, а структурами белого цвета, созревающими на свободных концах гиф гриба и напоминающими «муравьиные кольраби». Эти грибы нигде, кроме жилищ короедов, не растут. Однако, если из гнезда удалить самку, ухаживающую за грибным садом, то вскоре возделываемые грибы зарастают сорными грибами и погибают. Вслед за ними погибают от голода и личинки короеда. Такой своеобразный симбиоз обнаружен пока у небольшого количества видов из родов Anisandrus, Xyleborus, Xylotrupes, а также у некоторых ложнокороедов (Bostrychidae) и сверлил (Lymexylonidae).
Известны также комары родов Asphondylia, Lasioptera, Schizomia, Dasyneura, Contarinia, культивирующие грибы в галлах на зонтичных и бобовых растениях.
Между насекомыми и грибами также складываются взаимоотношения типа симбиоза, т. е. взаимовыгодного сожительства. Грибы дают насекомым богатую белками пищу, а взамен получают все необходимые для роста и развития условия. Здесь, как и в муравейниках или термитниках, налицо тесная взаимозависимость, невозможность раздельного существования ни насекомых, ни растений.
Роль травоядных животных в степных, луговых и тундровых биоценозах. Общий облик растительного покрова зависит не только от почвенно-климатических условий, но и от деятельности животных, находящихся в определенной связи с растениями. Изменение животного населения в любом из ландшафтов приводит к перемене в растительности. Полное же исключение животных сопровождается гибелью сообщества. Как отмечает Н. П. Наумов, в понятия «степь», «тундра», «лес», «луг» необходимо включать и животное население, и растительность потому, что они не только живут совместно, но и нуждаются друг в друге.
|
|
|
Широко известна роль слепушонков (Ellobius) в глинистых пустынях-такырах. Роясь в почве, они выбрасывают на поверхность холмики разрыхленной земли. На этих холмиках прорастают семена растений, которые в обычных условиях такыров не в состоянии пробиться через твердый слой засохшей глины. Постепенно растения покрывают некогда безжизненные пространства, создавая благоприятные условия для других растений и животных. Следовательно, слепушонки изменяют ландшафт, способствуют озеленению такыров.
Отмечаются случаи, когда истребление копытных в степях приводило к перерождению там растительности. Оказывается, многие злаки, основные степные растения, успешно развиваются и растут лишь при условии, если их объедают, «подстригают», копытные. Как только это исключается, они начинают вырождаться и в растительном сообществе происходит глубокая перестройка. Становится очевидным, что между копытными и злаками исторически сложились определенные межвидовые взаимоотношения. Благодаря им растения и животные приспособились к совместному существованию. На основе этого сосуществования и взаимного влияния и сформировался характерный степной биоценоз. Естественно, рассматривать копытных и злаки изолированно, вне связи с другими видами растений и животных, можно только условно, но доминирование взаимоотношений этих компонентов степного биогеоценоза не вызывает сомнения.
Примерно такую же роль играют копытные на полупустынных и пустынных пастбищах. Если они периодически не выедают полыней, то последние стареют и отмирают, а типичный ландшафт с определенным растительным покровом быстро перерождается. Такое явление характерно и по отношению к другим растительным ассоциациям. Повреждая растения неравномерно, избирательно, животные поддерживают тем самым нормальное соотношение их в сообществе.
В любом биогеоценозе между количеством и качеством животного населения и растительности должна быть определенная пропорция. Если увеличивается количество фитофагов, они уничтожают больше растений и в конце концов остаются без пищи, обрекая самих себя на гибель. С. С. Шварц, базируясь на экспериментальных данных, указывает, что «...максимальная продукция северных травостоев отмечается не тогда, когда она полностью ограждена от потребителей, а тогда, когда «пресс консументов» достигает определенной нормы. Лишь когда численность грызунов приближается к 1000 на гектар, продукция травостоя опускается ниже «спокойной нормы».
Следовательно, травоядные животные являются не разрушителями естественных фитоценозов, а их созидателями, неотъемлемой составной частью любого ландшафта. В природе исторически выработались и функционируют механизмы, поддерживающие наиболее выгодные количественные соотношения численности травоядных животных и растений, служащих им пищей.
Глава 10. ЗНАЧЕНИЕ БИОТИЧЕСКИХ ВЗАИМООТНОШЕНИЙ В РЕГУЛЯЦИИ ЧИСЛЕННОСТИ ВИДОВ
Общие черты конкуренции, хищничества и паразитизма как механизмов регуляции численности. Межвидовые взаимоотношения играют большую роль в динамике численности организмов в биоценозах. Хищники, уничтожая свои жертвы, влияют на численность последних. Такое же действие оказывают и паразиты. Ослабляя хозяев, они сдерживают их размножение, а наиболее угнетенные погибают. Однако наблюдения в природе показали, что нередко численность популяции хозяина из года в год остается более или менее стабильной и лишь незначительно колеблется у какого-то среднего уровня. Вместе с тем отмечаются и отклонения от этого правила, когда паразиты или хищники полностью подавляют популяцию жертвы, а затем сами начинают деградировать. Все это лишний раз подтверждает, что в природе имеется множество факторов, оказывающих влияние на естественное регулирование численности организмов в биоценозе и на взаимодействия между хищником и его добычей.
При конкурентных взаимоотношениях, хищничестве, паразитизме действуют генетические и экологические факторы. К. Уатт, анализируя полученные П. Эррингтоном данные наблюдений за взаимоотношениями норки (Mustella vison) и ондатры (Ondatra zibethica) пришел к выводу, что преследование ондатры хищником носит не случайный характер, что жертвами норки в первую очередь становятся те животные, которые представляют «биологический излишек» не только в экологическом, но π в генетическом смысле. Наиболее энергичные особи занимают самые удобные места обитания и активно защищаются от хищника, а ослабленные не способны активно противостоять ему. Хищничество, таким образом, приносит популяции жертвы двойную пользу. Оно прежде всего увеличивает поток энергии в биогеоценозе, приходящейся на одну жертву, и селективно элиминирует животных с низкой способностью добывать корм (медлительных, хилых, больных).
Математические модели Лотки – Вольтерры. Хищничество и сходное с ним явление паразитизма издавна вызывало у экологов большой интерес. Было предпринято много попыток разработать теорию взаимоотношений хищника и его добычи, паразита и хозяина, конкурирующих видов. Наибольшее признание получили работы A. Лотки (1923, 1925). Он один из первых предложил математическую модель взаимодействия животных в системе паразит – хозяин. Через год, независимо от Лотки, B. Вольтерра разработал математическую модель в системе хищник – жертва. Он же проанализировал и взаимоотношения между двумя конкурирующими видами.
Результаты исследований Лотки и Вольтерры в основном сходны. Это свидетельствует об отсутствии коренных различий в системах хищник – жертва и паразит – хозяин. Различия обнаружены лишь в их количественном соотношении, так как один хищник уничтожает много жертв, а паразитов может быть много на одного хозяина.
Сущность выводов Лотки сводится к тому, что истребление особей хозяина паразитами является функцией численности не только паразитов, но и хозяев. На основании этого он делает заключение, что той или иной численности хозяина соответствует определенная численность паразита и по мере возрастания плотности популяции хозяина увеличивается и плотность популяции паразита. Повышение же численности паразита приводит к снижению численности хозяина, а последнее опять снижает количество паразитов. И так волна за волной происходят периодические колебания численности популяций хозяина и паразита с небольшими отклонениями от какого-то оптимального уровня. Здесь действует динамическая саморегулирующаяся система, где волна подъема численности паразита следует за волной подъема численности хозяина. Модель взаимодействия популяций в системе паразит – хозяин полностью соответствует модели взаимодействия хищника и его жертвы, классический пример которой представлен на рис. 33.
Уравнения, предложенные Лоткой и Вольтеррой, можно использовать для моделирования конкурентных взаимоотношений в системах хищник – жертва, паразит–хозяин и др. Для некоторого ограниченного пространства, когда каждая из двух популяций имеет определенный уровень равновесия (К), одновременный рост их можно выразить следующим образом:
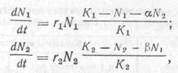
где N1 и N2 – соответственно численность жертвы и хищника; Κ1 и Κ2 – соответственно предельная плотность популяции жертвы и хищника, которая может быть достигнута при отсутствии конкурента; α и β – коэффициенты конкуренции, характеризующие тормозящее влияние второго вида на первый и наоборот (они, как правило, меньше единицы).
Из уравнений следует, что подавляющее влияние каждой особи второго вида на популяцию первого равно α/Κ1, а первого на популяцию второго – β/ Κ 2. Исход конкуренции зависит от относительных величин Κ1, К2, α и β. Различные их комбинации дают четыре возможных варианта воздействия популяций друг на друга (табл. 7).
Пользуясь приведенными уравнениями, можно определить, какой должна быть плотность каждой популяции, чтобы другая не имела возможности увеличивать свою численность. Очевидно, что N1 не может возрастать при N2 = Κ1/α, а когда Ν1 достигает величины Κ2/β, не может увеличиваться Ν2.
Таблица 7. Возможные варианты конкуренции, вытекающие из уравнений Лотки–Вольтерры (из Э. Пианка, 1981; с изменениями)
| Характер взаимоотношений | Варианты при условии, что | |
| вид 1 подавляет вид 2 (Κ2/β<Κ1) | вид 1 не подавляет вид 2 (Κ2/β> Κ1) | |
| Вид 2 подавляет вид 1 (Κ1/α < К2) | Каждый вид может выйти победителем (вариант 3) | Всегда побеждает вид 2 (вариант 2) |
| Вид 2 не подавляет вид 1 (Κ1/α > К2) | Всегда побеждает вид 1 (вариант 1) | Ни один вид не подавляет другой; устойчивое сосуществование (вариант 4) |
Для моделирования взаимоотношений типа хищник – жертва и паразит – хозяин Лотка и Вольтерра предложили уточненную пару уравнений:
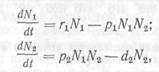
где Ν1 и Ν2 – плотности популяций соответственно жертвы и хищника; r 1и d2 – удельные скорости соответственно увеличения популяции жертвы и гибели популяции хищника; p1 и p2 – константы хищничества.
Из этих уравнений следует, что в отсутствие хищника популяция жертвы растет экспоненциально с потенциально неограниченной скоростью. Произведение N 1 N2 отражает число контактов между двумя видами. Умножив его на p2, получим максимальную скорость увеличения популяции хищника (p 2 N 1 N 2 ), а на р 1 - скорость роста популяции жертвы (p 1 N 1 N 2 )/
Пользуясь этими уравнениями, можно также определить условия, при которых устанавливается равновесие между плотностями популяций жертвы N 1и хищника N2, т. е. система хищник – жертва становится стабильной. Для этого приравняем нулю производные по времени от N 1и N2. В результате получим:
r 1 N 1 -p 1 N 1 N2=0,
p2N 1 N2-d2N2 =0.
Отсюда следует, что стабильная плотность популяции жертвы (N 1 = d2/p2) может наступить только тогда, когда плотность популяции хищника установится на уровне N2=rl/pl.
В результате своих наблюдений Вольтерра сформулировал три закона.
Закон периодического цикла: колебания численности двух видов являются периодическими и зависят от коэффициента роста популяций хищника и жертвы и исходной относительной численности.
Закон сохранения средних величин: средняя численность популяции обоих видов остается постоянной, независимо от первоначальной численности до тех пор, пока скорости увеличения и уменьшения популяций, а также интенсивность хищничества постоянны.
Закон нарушения средних величин: если уничтожать особей обоих видов в одинаковой степени (пропорционально плотности их популяций), то средняя численность популяции жертвы будет расти, а хищника · – падать.
Модели взаимоотношений хищника (паразита) и жертвы (хозяина), созданные Лоткой и Вольтеррой, подвергались критике, поскольку их основу составляют упрощенные случаи, не встречающиеся в природе. Тем не менее разработанные ими положения имеют большое теоретическое и практическое значение. Они показывают, как при постоянных условиях взаимоотношения хищников и добычи сами по себе могут возбудить численные колебания обоих. Это происходит потому, что у хищника вероятность поймать жертву больше при увеличении плотности, чем при возрастании численности ее.
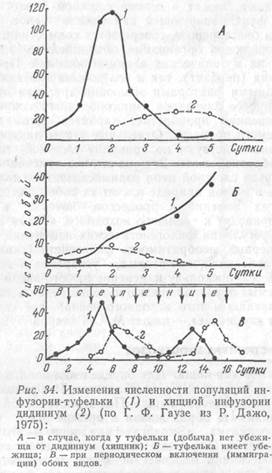
Опыты Г. Ф. Гаузе. Гаузе, решив проверить положения Лотки – Вольтерры, в 1930–1932 гг. провел эксперименты в системе, включающей две популяции – инфузории Paramaecium bursaria (хищник) и дрожжей Saccharomyces pombe (жертва), и также обнаружил циклы колебаний численности как одной, так и другой. Подобные результаты получены с популяциями инфузории-туфельки (Paramaecium aurelia) и ее добычи – дрожжей Saccharomyces exiguus. Однако дальнейшие исследования взаимоотношений хищной инфузории Didinium с инфузорией Paramaecium показали, что хищник может полностью уничтожить свою добычу, но после этого сам погибает от недостатка пищи. Если же для жертвы создаются какие-либо убежища, где могут спрятаться отдельные особи ее, то через определенное время популяция хищника погибает от голода, а сохранившиеся индивидуумы парамеции начинают интенсивно размножаться. Эти опыты свидетельствовали, что устойчивое взаимодействие хищника и жертвы проявляется чрезвычайно редко и только тогда, когда отдельные особи последней недоступны для хищника или в случае пополнения ее популяции пришельцами со стороны, т. е. иммигрантами.
Анализируя конкурентные взаимоотношения простейших, Гаузе обосновал положение, названное «принципом Гаузе» (теорема Гаузе). Это положение получило широкое распространение у нас и за рубежом. Оно сводится к следующему: 1) подавляющий популяцию эффект сильнее сказывается на популяции другого вида, чем на собственной; 2) в результате конкуренции истребляется тот вид, которому наносится больший вред; 3) подавляющий популяцию эффект слабее сказывается на популяции другого, чем на собственной. В последнем случае, когда каждому из видов свойственна определенная экологическая ниша с более благоприятными именно для него условиями, складывается ситуация, при которой популяции обоих видов могут сосуществовать неопределенно долгое время (рис. 34).
В природе взаимоотношения конкурирующих видов, взаимодействия популяций в системе хищник (паразит) – жертва (хозяин) в чистом виде никогда не наблюдаются. Хищник, к примеру, питается не одним видом, жертва также преследуется многими хищниками. Кроме того, все взаимоотношения складываются в конкретных условиях, на которые каждый член системы реагирует по своему. Поэтому строго циклических колебаний численности хищников (паразитов) и добычи (хозяина), предполагаемых математической теорией Лотки – Вольтерры, Гаузе и других исследователей, в природе не наблюдается. Тем не менее принцип математического моделирования сложных межвидовых взаимоотношений - позволяет значительно углубить наши представления о механизмах регуляции численности организмов в биоценозах. Н. П. Наумов в связи с этим отмечает: «Динамика населения любого вида представляет собой результат взаимодействия со всеми условиями существования. Среди последних роль ведущего фактора деятельность хищников может играть только при определенной ситуации. Знание и умение создавать ее имеет большое практическое значение, так как на нем основан так называемый биологический способ борьбы с вредителями».
Конкуренция, хищничество, паразитизм и другие биотические связи, возникающие в биоценозе между различными видами, лежат в основе сложной биологической системы, функционирующей как единое целое. В этой системе, в биогеоценозе, совершается совместная, сопряженная, эволюция организмов, объединенных на базе трофических и топических взаимоотношений. При этом как хищник (паразит), так и его добыча (хозяин) являются мощными факторами эволюции друг для друга, а направление ее благодаря многообразным коррелятивным отношениям приобретает характер автоматически регулируемого процесса. Отдельные этапы данного процесса взаимодействуют по принципу сложной положительной обратной связи. Это значит, что изменение одного из звеньев сложной цепи взаимосвязанных явлений и процессов в живой природе влечет за собой ряд изменений других явлений и процессов, которые в свою очередь приводят к усилению исходного. «Такая автоматическая регуляция филогенетических изменений объясняет, во-первых, необратимость филогенетических изменений организации... а во-вторых,– ускорение эволюционного процесса, поскольку изменение любого звена в цепи регулируемых обратной связью явлений ведет к дальнейшему усилению и того исходного звена, где впервые возникает изменение»,– пишет М. С. Гиляров.
Глава 11. СТРУКТУРА БИОЦЕНОЗОВ
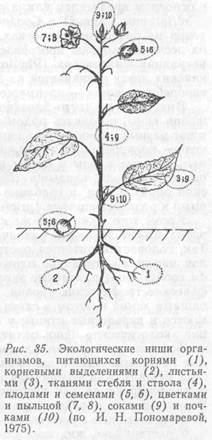
Экологическая ниша. Для определения роли, которую играет тот или иной вид в биогеоценозе, И. Гринелл ввел понятие «экологическая ниша». Под экологической нишей следует понимать образ жизни и прежде всего способ питания организма. «Экологическая ниша» – это абстрактное понятие. Оно включает химические, физические, физиологические и биотические факторы, необходимые организму для жизни, и определяется его морфологической приспособленностью, физиологическими реакциями и поведением. По образному выражению Ю. Одума, термин «экологическая ниша» относится к роли, которую играет организм в экосистеме, иначе говоря, местообитание – это «адрес», тогда как ниша – «профессия» вида. Чтобы охарактеризовать экологическую нишу вида, необходимо знать, чем он питается и кто его поедает, способен ли он к передвижению и, наконец, как он воздействует на другие элементы биогеоценоза (рис. 35).
|
В разных частях света и на разных территориях встречаются неодинаковые в систематическом отношении, но экологически сходные виды. Например, травянистая или лесная растительность Австралии по видовому составу существенно отличается от растительности сходного климатического района Европы или Азии, но как продуценты в своих биогеоценозах они выполняют аналогичную функцию, т. е. занимают в принципе одни и те же экологические ниши. То же можно сказать и о сосне в борах и дубе в дубравах. Это касается также антилоп Африки и кенгуру Австралии. Сумчатый волк (Thylacinus cynocephalus) в Тасмании и обыкновенный волк в Европе тоже ведут одинаковый образ жизни, обладают сходным типом питания, т. е. в различных биогеоценозах они выполняют одинаковую функцию.
Нередко бывает и наоборот – одному виду в разных биоценозах свойственны различные экологические ниши. Чаще всего это связано с доступностью пищи и наличием конкурентов. Особенно много примеров подобного рода среди насекомых. Короед-типограф (Ipstypographus) в Сибири и европейской части СССР является в основном вредителем ели, а на Кавказе – сосны. Прус (Culliptamus italicus) в Средней Азии повреждает растения на орошаемых участках, а в Западной Сибири на песчаных хорошо прогреваемых солнцем местах. Виноградная филлоксера (Phylloxera vastatrix) на американских лозах развивается на листьях и на корнях, а на европейских лозах – преимущественно на корнях.
Вместе с тем часто близкие в систематическом отношении виды, поселяясь рядом в одном биоценозе, занимают разные экологические пиши. Это приводит к уменьшению конкурентного напряжения между ними. Например, водяные клопы в одном и том же водоеме играют различную роль: гладыши (Notonecta) ведут хищнический образ жизни, а гребляки (Corixa) питаются мертвыми и разлагающимися организмами.
Кроме того, один и тот же вид в разные периоды развития может занимать различные экологические ниши. Так, головастик питается растительной пищей, а взрослая лягушка – типичное плотоядное животное, поэтому им свойственны различные экологические ниши и специфические трофические уровни. Разными экологическими нишами зимой и летом в связи с миграциями характеризуются и перелетные птицы: у насекомоядных птиц зимние экологические ниши отличаются от летних. Гусеницы бабочек, личинки пилильщиков, мух, жуков и взрослые насекомые также занимают различные экологические ниши. Среди водорослей имеются виды, которые функционируют то как автотрофы, то как гетеротрофы. В результате в определенные периоды жизни они занимают те или иные экологические ниши.
У растений же, живущих в одном ярусе, экологические ниши сходны. Именно это ослабляет конкуренцию между растениями разных ярусов и обусловливает освоение ими различных экологических ниш. В биоценозе разные виды растений занимают различные экологические ниши, что ослабляет межвидовую конкурентную напряженность. Однако один и тот же вид в различных природных зонах может занимать разные экологические ниши. Например, сосна (Pinus silvestris) и черника (Vaccinium myriillus) в бору-черничнике, водные растения рдесты (Potamogeton), кубышка (Nuphar luteum), водокрас (Hydrocharius morsus), ряски (Lemnaceae) поселяются вместе, но распределяются по различным нишам. Черника и седмичник (Trientalis europaea) в лесах умеренной полосы являются типичными теневыми формами, а в тундре и лесотундре растут на открытых пространствах и становятся световыми.
|
|
|
|
|
Дата добавления: 2014-11-06; Просмотров: 1013; Нарушение авторских прав?; Мы поможем в написании вашей работы!