КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Экология сообществ и экосистем 5 страница
 |
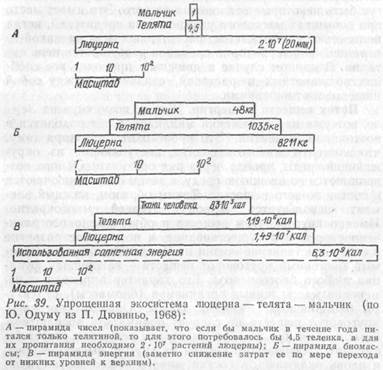
Правило пирамиды чисел универсально и объективно отражает круговорот веществ и поток энергии в биосфере. В масштабе всей биосферы это правило никогда не нарушается. Правда, на незначительных участках могут быть некоторые отклонения от него. Это имеет место при вспышках массового размножения вредителей, когда полностью уничтожается растительность и на какой-то ограниченной территории временно разрушается цепь питания. В данном случае в движение приходит все сообщество животных и растений, связанных между собой пищевыми отношениями.
Поток веществ и энергии. Жизнь, возникнув на Земле, вот уже на протяжении миллиардов лет находится в постоянном развитии. Это происходит благодаря тому, что элементы живого вещества, поступающие из окружающей среды, пройдя через ряд организмов, снова возвращаются во внешнюю среду, а затем опять включаются в состав живого вещества. Таким образом, каждый элемент используется живой материей многократно. Именно круговоротом веществ и обусловлено неограниченное временем существование и постоянное развитие и совершенствование жизни на Земле. Этот так называемый биогенный круговорот веществ – важнейшая функция любого биогеоценоза. Его характер определяют изменения массы живых организмов (биомассы), структуры биогеоценоза, химизма среды. Однако биогенный круговорот веществ не следует понимать в абсолютном смысле. Как бы там ни было, эти вещества, переходя с одного трофического уровня на другой, высвобождаясь и вновь включаясь в состав живого вещества, частично исключаются из круговорота. В результате на Земле происходит накопление органических соединений в виде залежей полезных ископаемых (торф, уголь, нефть, газ, горючие сланцы). Но все это не отвергает общего правила. Существенно биомасса на Земле не накапливается, а удерживается на каком-то определенном уровне, поскольку она постоянно разрушается и вновь созидается из одного и того же строительного материала, т. е. в ее пределах протекает беспрерывный круговорот веществ.
|
|
|
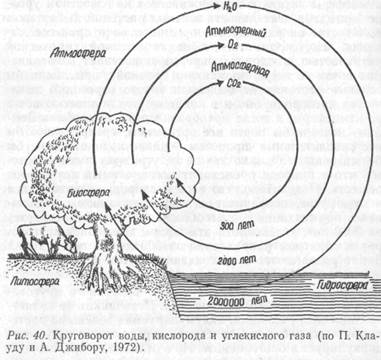
Биогенный круговорот веществ принял определенный характер с появлением зеленых растений, осуществляющих процессы фотосинтеза. Рассмотрим это на примере круговорота кислорода – продукта фотосинтеза растений. Практически весь молекулярный кислород земной атмосферы возник и поддерживается на известном уровне благодаря деятельности зеленых растений. В большом количестве он расходуется организмами в процессе дыхания. Но, кроме того, обладая высокой химической активностью, кислород непрерывно вступает в соединения почти со всеми элементами земной коры. Если бы зеленые растения не выделяли такого огромного количества кислорода, он бы в конце концов полностью исчез из атмосферы, и тогда преобразился бы весь облик Земли – исчезли бы почти все организмы, прекратились бы все окислительные процессы – планета наша стала бы безжизненной. Однако это ей не угрожает именно потому, что в природе происходит нескончаемый круговорот веществ. Подсчитано, что весь кислород, содержащийся в атмосфере, оборачивается через организмы (связываясь при дыхании и высвобождаясь при фотосинтезе) за 2000 лет, углекислота атмосферы совершает круговорот в обратном направлении за 300 лет, а все воды на Земле разлагаются и воссоздаются путем фотосинтеза и дыхания за 2000000 лет (рис. 40).
Однако для столь грандиозного биогенного круговорота веществ необходима энергия. Источником ее является солнечная радиация, аккумулируемая зелеными растениями-автотрофами. Солнечная энергия также регулярно циркулирует в биогеоценозе. Но в отличие от круговорота веществ, который протекает по замкнутому кругу, переходя с одного трофического уровня на другой, она постоянно расходуется. До 30 % ее рассеивается в атмосфере или отражается облаками и поверхностью Земли, до 20 % поглощается в верхних слоях облаками (водяные пары, капельки воды и пылевые частицы), приблизительно 50 % достигает суши или поверхности океана и поглощается в форме тепла. Лишь ничтожная часть, всего около 0,1–0,2 % энергии, получаемой Землей от Солнца, улавливается зелеными растениями и обеспечивает весь биогенный круговорот веществ в биосфере.
|
|
|
Более половины энергии, связанной при фотосинтезе, тут же расходуется на дыхание растений, а остальная поступает в пищевые цепи.
Таким образом, энергия Солнца, утилизированная зелеными растениями, превращается в потенциальную энергию химических связей органических соединений, из которых строится само тело растений.
В организме растительноядного животного эти органические вещества окисляются с выделением такого количества энергии, которое было затрачено на их синтез растением. Часть ее используется для жизни животного, а остальная, согласно второму закону термодинамики (переход энергии из одной формы в другую сопровождается снижением количества полезной энергии), превращается в тепло и рассеивается в пространстве, т. е. уходит из биоценоза (энтропия).
Поток энергии в экосистеме может быть проиллюстрирован схемой простой цепи питания (рис. 41). Солнечная энергия, полученная растением, лишь частично используется в процессе фотосинтеза углеводов. Фиксированная в углеводах энергия представляет собой валовую продукцию биогеоценоза (П в).
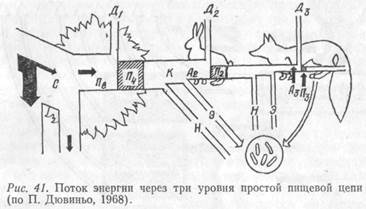
Углеводы идут на построение протоплазмы и рост растений; причем часть их энергии затрачивается на дыхание (Д1). В результате чистая продукция (П ч) определяется по формуле:
Пч = Пв - Д1
Таким образом, поток энергии, проходящий через уровень продуцентов, т. е. валовую продукцию, можно представить так:
Пв = Пч + Д1
Определенное количество созданных продуцентами веществ служит кормом (К) фитофагов, остальное в конце концов отмирает и перерабатывается редуцентами (Н). Корм, ассимилированный фитофагами (А2), лишь частично используется для образования их биомассы (П2). В основном он растрачивается на обеспечение энергией процессов дыхания (Д2) и в некоторой степени выводится из организма в виде выделений и экскрементов (Э). Поток энергии, проходящий через второй трофический уровень, выражается следующим образом:
|
|
|
А2 = П2 + Д2.
Консументы второго порядка (хищники) не истребляют всю биомассу своих жертв, но и из того количества ее, которое они уничтожают, лишь часть используется на создание биомассы их собственного трофического уровня.
Остальная же часть в основном затрачивается на энергию дыхания, а также выделяется с экскретами и экскрементами. Поток энергии, проходящий через уровень консументов второго порядка (плотоядные), выражается формулой:
А3 = П3+Д3.
Анализируемая схема наглядно показывает, что поток энергии, который выражается количеством ассимилированного по цепи питания вещества, на каждом трофическом уровне уменьшается, т. е.
ПЧ>П2>П3 и т. д.
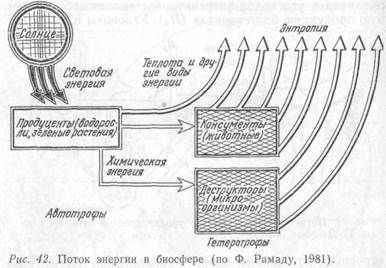
Таким образом, поскольку определенное количество вещества может быть использовано каждым биоценозом неоднократно, а порция энергии лишь один раз, правильнее говорить, что в экосистеме происходит каскадный перенос энергии (рис. 42) и круговорот веществ.
Биологическая продуктивность. Биологическая продуктивность – это воспроизведение биомассы растений, микроорганизмов и животных, входящих в состав биогеоценоза. Этот процесс протекает в природе с определенной скоростью. Поэтому биологическую продуктивность можно выразить продукцией за сезон, за год, за несколько лет или за любую другую единицу времени. Для наземных и донных организмов она определяется количеством биомассы на единицу площади, а для планктонных и почвенных – на единицу объема.
Следовательно, биологическая продуктивность представляет собой количество воспроизведенной биомассы на 1 кв. м площади (или на 1 куб. м объема) в единицу времени и выражается чаще всего в граммах углерода или сухого органического вещества. Биологическую продуктивность нельзя смешивать с биомассой. Допустим, за год планктонные водоросли на единицу площади синтезируют столько же органического вещества, сколько и высокопродуктивные леса, однако биомасса последних в сотни тысяч раз больше.
|
|
|
Биомасса того или иного биоценоза не дает четкого представления о его продуктивности. Это связано с тем, что скорость образования биомассы (продуктивность) в разных биоценозах неодинакова. Поэтому биоценозы различаются не только биомассой, но и продуктивностью, т. е. скоростью создания определенного количества биомассы. Луговые степи дают больший годовой прирост биомассы, чем хвойные леса. При средней фитомассе 23 т/га годовая продукция их составляет 10 т/га, тогда как у хвойных лесов при фитомассе 200 т/га она равна 6 т/га. Популяции мелких млекопитающих по сравнению с крупными обладают большой скоростью роста и размножения и дают более высокую продукцию при равной биомассе.
Таким образом, чтобы оценить значение вида (группы видов) для круговорота веществ и в отношении его биологической продуктивности в биоценозе или в биогеоценозе в целом, нужно знать не только его биомассу, но и относительную скорость прироста или время ее полного возобновления.
Продукция каждой популяции за определенное время представляет собой сумму прироста всех особей, включая отделившиеся от организма образования и устраненные (элиминация) по разным причинам особи (смерть, миграция). В том случае, когда все особи доживают до конца изучаемого периода, продукция равна приросту биомассы. В природе это исключено, и продукция популяции обычно рассчитывается по формуле:
Р=(В2-В1)+Е,
где Ρ – продукция; В1 и В2- –соответственно начальная и конечная биомасса; Ε – элиминация.
Это так называемая чистая продукция. Валовая продукция включает в себя прирост (чистая продукция) и затраты на энергетический обмен.
Необходимо различать первичную продукцию, т. е. продукцию автотрофных организмов, и первичную продуктивность, т. е. скорость, с которой автотрофные организмы (продуценты) в процессе фотосинтеза связывают энергию и запасают ее в форме органического вещества.
Подсчитано, что солнечная энергия, достигающая поверхности Земли в течение года, исчисляется в 5-1020 ккал (21-1020 кДж). Это составляет 9 млрд. ккал (37,8·109 кДж) на гектар. Один гектар леса в средних широтах продуцирует до 6 т древесины и 4 т листьев, сжигание которых дает 46 млн. ккал (193,2-106 кДж). Значит, эффективность первичной продуктивности леса, т. е. эффективность использования растениями солнечной энергии для создания органического вещества, составляет всего около 0,5 % (46X100:9). Конечно, цифры эти чрезвычайно относительны, поскольку эффективность первичной продуктивности зависит от возраста леса, количества деревьев, погодных условий и многих других факторов. Но тем не менее они дают представление о коэффициенте полезного действия биоценоза.
Консументы образуют свою биомассу. Для обозначения биомассы и скорости ее образования консументами применяются термины «вторичная продукция», т. е. продукция гетеротрофных организмов, и «вторичная продуктивность», т. е. скорость образования продукции гетеротрофами. Как уже отмечалось, поток энергии от продуцентов к консументам сопровождается потерями ее. Дело в том, что значительная часть съеденного гетеротрофами корма расходуется на теплопродукцию, на выработку энергии, необходимой для их жизнедеятельности, и лишь небольшое количество его (1,3–2 %) используется на создание вторичной продукции. Например, для получения 1 кг говядины требуется от 70 до 90 кг свежей травы.
При этом необходимо учитывать, что все виды, дающие вторичную продукцию, возникают на основе утилизации вещества и энергии первичной продукции. Но так как при переходе с одного трофического уровня на другой энергия частично затрачивается на нужды энергетического обмена и рассеивается, то продукция каждого последующего трофического уровня меньше продукции предыдущего. Например, продукция (ее выход) фитофагов всегда больше, чем у живущих за их счет хищников.
Большое значение в механизме биологического продуцирования имеют гетеротрофные организмы, утилизирующие поступающее со всех трофических уровней мертвое органическое вещество, частично минерализуя его, частично превращая в вещество микробных тел. Последнее служит важным источником питания многих водных и почвенных животных.
Кроме первичной и вторичной продукции биоценозов, различают промежуточную и конечную продукцию. Промежуточная продукция отличается тем, что после потребления другими членами биогеоценоза возвращается в круговорот веществ этой же системы. Конечная продукция исключается из данного биогеоценоза, т. е. выводится за его пределы. Это, к примеру, продукция, получаемая человеком в процессе возделывания сельскохозяйственных культур, разведения домашних животных, охоты, промысла и т. д.
Глава 13. ДИНАМИКА И СТАБИЛЬНОСТЬ ЭКОСИСТЕМ
Суточная и годовая динамика экосистем. Одной из характерных особенностей биоценозов является их суточная и годовая динамика. В каждом естественном биоценозе имеются группы организмов, активность жизни которых выпадает на разное время суток. Одни активны в ночное время и днем обычно скрываются в каких-либо убежищах. Другие, наоборот, пассивны ночью. В результате в составе и в соотношении отдельных видов биоценоза происходят периодические изменения, поскольку отдельные организмы на определенное время выключаются из него. При этом суточную динамику биоценоза обеспечивает не только животное, но и растительное население. У растений в течение суток также изменяются интенсивность и характер физиологических процессов, в ночные часы не происходит фотосинтез, у ряда растений цветки раскрываются только ночью и опыляются ночными животными, другие же приспособлены к опылению днем.
Большое значение в динамике биоценозов имеют суточные миграции животных. Они зависят от изменения характера действия различных факторов в течение суток. Особенно типичны суточные миграции морского планктона. Представители зоо- и фитопланктона в Каспийском море, например, днем держатся на глубине от 100 до 350 м, а ночью поднимаются в поверхностные слои. Вертикальные суточные миграции свойственны и почвенным обитателям.
Суточная динамика биогеоценозов в основном связана с ритмикой природных явлений и носит строго периодический характер.
Но, кроме этого, могут происходить и непериодические изменения активности и численности тех или иных компонентов экосистемы в течение суток, связанные с действием нерегулярных факторов среды. Так, сильные дожди или засухи приводят к перемещению животных, изменению их активности, влияют на интенсивность некоторых жизненных процессов у растений. Во время сильных ливней даже морской планктон из-за опреснения поверхностных слоев воды мигрирует в глубину.
Более существенные отклонения в биоценозах наблюдаются при сезонной динамике. Они обусловлены биологическими циклами организмов, зависящими от сезонной цикличности природных явлений. Смена времен года оказывает значительное влияние на жизнедеятельность растений и животных (периоды цветения, плодоношения, активного роста, листопада и зимнего покоя у растений; спячка, зимний сон, диапауза и миграции у животных).
В разных широтах длительность биологических сезонов неодинакова. Поэтому сезонная динамика биогеоценозов арктической, умеренной и тропической зон различна. Наиболее четко она выражена в экосистемах умеренного климата и в северных широтах.
В связи с тем что характер суточных и сезонных изменений более или менее постоянен в течение длительного времени (столетия и даже тысячелетия), в биоценозах исторически сформировались механизмы, приводящие сообщество в целом в соответствие с периодикой изменения условий обитания.
Экологические сукцессии. В процессе суточной и сезонной динамики целостность биоценозов обычно не нарушается. Биоценоз испытывает лишь периодические колебания качественных и количественных характеристик. Но он может подвергаться и воздействию сил, которые существенно изменяют или полностью разрушают сообщество. В таких случаях развивается другой, более приспособленный к новым условиям биоценоз. Подобная последовательная смена одного биоценоза другим называется экологической сукцессией (succession – последовательность, смена). В целом цепь сменяющих друг друга биоценозов называется сукцессионным рядом или серией (сериальные стадии). В сукцессионном ряду каждый биоценоз представляет собой определенную стадию формирования конечного, завершающего, или так называемого климаксного сообщества. По мере усложнения сообщества усложняются и связи между популяциями. Менее приспособленные к новым условиям замещаются более приспособленными, и так до тех пор, пока не появятся виды, которых условия среды полностью устраивают и они уже не замещаются другими видами. В результате сообщество становится стабильным и достигает своей завершающей стадии.
Концепция о заключительной, завершающей фазе развития биоценоза, точнее биогеоценоза, как учение о климаксе имеет некоторые негативные стороны. Дело в том, что в любом биогеоценозе действуют определенные противоречивые движущие силы развития. Различают противоречия между синтезирующими и разрушающими органические вещества группами организмов (внутренние противоречия) и противоречия между биоценозом и опосредствованной им средой обитания (внешние противоречия). Единство этих двух типов противоречий лежит в основе существования и развития биоценозов на Земле. Поэтому сами биоценозы также относительно устойчивы. Выражается эта относительность и противоречивость в колебаниях численности отдельных популяций, особенно в периоды вспышек массового размножения, во время вторжения новых, устойчивых видов и т. д. В результате могут нарушаться биоценотические связи и происходить заметные, иногда необратимые, изменения в строении биоценозов.
С позиций диалектического материализма существование абсолютно климаксного биоценоза невозможно, так как взаимное воздействие компонентов его никогда не прекращается, а следовательно, и не останавливается биогеоценотический процесс. В результате совершаются сукцессии. Этот процесс постоянен, ибо развитие биоценозов, как и любого природного явления, не может остановиться. Со временем микроорганизмы, растения и животные в биогеоценозе (его составная живая часть – биоценоз) приобретают некоторую относительную устойчивость – динамическое равновесие, сбалансированность отдельных компонентов в сообществе. По такое климаксное состояние биоценоза не бывает постоянным во времени и пространстве, раз и навсегда установившимся. Поэтому термин «климакс» можно использовать лишь условно в смысле приобретения биоценозом определенной замедленности в развитии, поскольку даже при кажущейся неизменности в биоценозе все-таки протекают процессы, подготавливающие его к сукцессии.
В историческом разрезе смена фауны и флоры по геологическом периодам не что иное, как смена сообществ, замена одного типа биоценоза другим, т. е. – экологическая сукцессия. Эти сукцессии тесно связаны с геологическим и климатическими изменениями, а также с эволюцией видов.
Нередко сукцессии совершаются в сравнительно короткие промежутки времени и тогда они не связаны с эволюцией видов. Ведь биоценоз в течение ряда лет не остается постоянным. Изменяются условия жизни, исчезают одни и появляются другие группы организмов, меняются связи между популяциями, а вслед за этим – и биоценоз в целом. Иногда такие изменения происходят очень медленно, даже столетиями. А бывает, что и относительно быстро. Лесной пожар может моментально уничтожить сложившийся на протяжении тысячелетий устойчивый биоценоз, и на пожарище начинает довольно быстро создаваться новое сообщество. При этом будет наблюдаться частая смена ряда сообществ. В конечном итоге устойчиво восстанавливается биоценоз исходного типа (или же он незначительно отличается от первоначального). Классическим примером сукцессии с образованием устойчивого биоценоза является зарастание озера и возникновение на его месте торфяного болота или формирование елового леса на брошенных землях (рис.43).
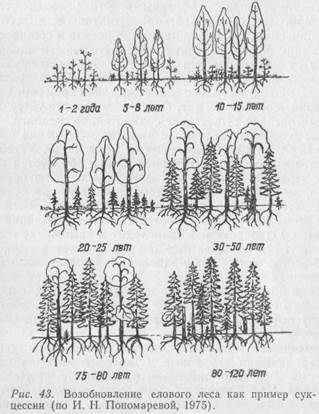
Еловый лес в своем развитии проходит несколько этапов. Первыми на заброшенной пашне из древесных пород появляются береза, осина и ольха, поскольку семена этих деревьев легко разносятся ветром. Попав на слабо задернованную почву, они прорастают. Такие первопоселенцы обычно называются пионерами. Наиболее стойкие из них заселяют заброшенную или распаханную территорию, утверждаются там и постепенно изменяют среду, создавая новые условия, к которым сами со временем оказываются неприспособленными. Эти условия становятся пригодными для растений-захватчиков, вытесняющих пионеров и начинающих доминировать в сообществе до тех пор, пока в результате их деятельности вновь не изменятся условия и они не начнут замещаться более приспособленными формами. Условия, благоприятные для ели, создаются только после смыкания крон берез, т. е. примерно через 30–50 лет. Постепенно формируется смешанный лес. Он существует сравнительно недолго, так как светолюбивые березы не выносят затемнения и под пологом елей их возобновления не происходит. Устойчивый еловый лес на заброшенной пашне образуется примерно через 80–120 лет после первых всходов березы.
В процессе развития березняков, ольховников, осинников, а затем и елового леса в биоценоз включаются все новые виды растений и животных, сопровождающие доминантов. Одновременно происходит и замещение одних видов другими. По мере того как увеличивается их число, в сообществе возникают и заполняются новые экологические ниши.
Таким образом, сукцессия протекает как медленное и в какой-то степени случайное замещение одних популяций другими, а не путем резкой, скачкообразной смены сообществ.
Первичные и вторичные сукцессии. Различают сукцессии первичные и вторичные. Сукцессия, которая начинается на абсолютно лишенном жизни месте, называется первичной. При первичных сукцессиях скорость изменения сообществ, как правило, невелика. Сукцессионные ряды, последовательно сменяющие друг друга, удерживаются значительный промежуток времени, а достижение биоценозом климаксного состояния затягивается иногда на столетия и больше. К типичным первичным сукцессионным образованиям биоценозов в масштабе геологических периодов относится формирование фауны и флоры на вновь появившейся песчаной дюне, на лавовом потоке, в послеледниковый период, после поднятия материков и т. д.
Если сообщество развивается на месте, где ранее существовал хорошо развитый биоценоз, то сукцессия будет вторичной. В таких местах обычно сохраняются богатые жизненные ресурсы. Поэтому вторичные сукцессии приводят к образованию климаксного сообщества значительно быстрей, чем первичные. В современных условиях вторичные сукцессии наблюдаются повсеместно. Они обусловлены последствиями, возникающими в результате пожаров, наводнений, распашки степей, вырубки лесов, осушения болот.
Протекают сукцессии и внутри биоценозов. Примером быстрых сукцессии комплексов организмов местного характера на мелких участках являются дупла деревьев. Дупла обладают сравнительно устойчивым микроклиматом. От величины дупла, толщины его стенок, состояния древесины зависят условия, благоприятствующие заселению его теми или иными видами организмов. Постепенно, в процессе сукцессии, в дупле формируется своеобразный биоценоз. При заселении дупла каким-либо видом птиц или млекопитающих сообщество мелких организмов претерпевает коренную перестройку. Вокруг основного обитателя дупла концентрируются сопутствующие виды (квартиранты, паразиты, симбионты, комменсалы). Между хозяином дупла и остальными жителями его устанавливаются строго определенные количественные пропорции. В результате на какой-то период образуется своеобразное климаксное сообщество.
Еще более характерные сукцессии протекают в норах. Лишенная обитателей нора имеет температуру и влажность, близкие к окружающей среде. Если же нора обитаема, то в ней создается вполне специфический микроклимат. Норовые биоценозы нередко насчитывают до 200 и более квартирантов. Многие из них связаны с хозяином норы не только пространственными, но и пищевыми взаимоотношениями (паразиты). Естественно, что устойчивый норовый биоценоз развивается постепенно, в процессе сменяющих друг друга сукцессионных рядов.
Хорошо изучены сукцессии в разлагающихся стволах деревьев. В течение первых 3–4 лет древесину заселяют в основном ксилофаги из насекомых и деревоокрашивающие грибы. Позже появляются дереворазрушающие грибы, увеличивается количество ксилофагов, хищных насекомых. Затем в гниющей древесине поселяются сапроксилофаги, т. е. насекомые, питающиеся гнилой древесиной. Вместе с этим растет численность хищников и изменяется их видовой состав. На 8–10-м году количество ксилофагов начинает сокращаться, соответственно снижается и число хищников, а число сапроксилофагов возрастает. Затем появляются шляпочные грибы. Примерно через 10–15 лет древесина полностью разрушается и специфичный биоценоз исчезает.
Интересна динамика биоценозов в слое растительного опада, при обрастании камней, заселении трупов животных, высыхающего навоза. Например, трупы животных на различных стадиях разложения заселяются разными комплексами организмов. Из насекомых вначале появляются могильщики и мертвоеды (Necrophorus, Silpha), затем мухи (Calliphoridae, Muscidae, Sarcophagidae) и жуки-карапузики (Histeridae). Уже высохшие трупы заселяются кожеедами (Dermestidae).
Известны сукцессии биоценозов плодовых деревьев по мере изменения их состояния. Молодые неплодоносящие сады заселяются многоядными насекомыми-фитофагами, среди которых наиболее характерны хрущи (Melolonthinае) и чернотелки (Tenebrionidae); из бабочек – зимняя пяденица (Operoptera brumata), кольчатый шелкопряд (Malacosoma neustria), различные листовертки (Tortricidae). Кроме этого, многочисленны тли (Aphidodea) и щитовки (Diaspidae). Когда сад начинает плодоносить, появляются насекомые, питающиеся цветками и плодами (яблонная и другие плодожорки – Laspeyresia, Grapholitha, яблонный цветоед – Anthonomus pomorum, вишневый слоник – Rhynchites auratus, казарка – R. bacchus). По мере старения сада в нем обнаруживаются стволовые вредители (короеды – Ipidae, древоточцы – Cossidae, стеклянницы – Aegeriidae). Численность их постоянно растет. Вслед за изменением фитофагов меняется в количественном и качественном отношении и фауна паразитов и хищников.
Общие закономерности сукцессии. Экологическая сукцессия является результатом изменений, которые вносятся в среду обитания самими сообществами. Это закономерный, направленный процесс. Он заканчивается образованием климаксного биоценоза, характеризующегося максимальной величиной биомассы и наибольшим разнообразием слагающих его видов. Климаксный биоценоз находится в состоянии гомеостаза.
Общие закономерности сукцессии, согласно Ю. Одуму, сводятся к следующему. В направлении сукцессии от первичных стадий к климаксу, или от юных к стареющим сообществам, изменяется видовая и органическая структура, а также миграция энергии – метаболизм сообщества.
Видовой состав в сукцессионном ряду меняется сначала быстро, а затем более медленно. Видовая численность автотрофов увеличивается в первичных и часто в начале вторичных сукцессии, хотя на стадиях старения может снижаться. Количество видов гетеротрофов растет до относительно поздней стадии. Таким образом, видовое разнообразие поначалу увеличивается, потом стабилизируется и в итоге, на стадиях старения, снижается.
При изменении органической структуры сукцессии увеличивается общая биомасса сообщества и количество мертвого органического вещества, в течение ранней фазы первичной сукцессии возрастает количество хлорофилла.
Метаболизм (миграция энергии) по ходу смены сообществ также изменяется. Связи по линии пищевой цепи становятся все более сложными. Общая продуктивность повышается в течение ранней фазы первичной сукцессии и мало или совсем не изменяется во время вторичной сукцессии. Чистая продуктивность сообщества снижается, а дыхание его растет.
Ю. Одум указывает, что наибольшее значение в перечисленных закономерностях имеют 4 аспекта.
1. Видовой состав растений и животных в процессе сукцессии непрерывно меняется. Это свидетельствует о том, что виды, доминирующие на начальных стадиях сукцессии, по мере достижения биоценозом климаксного состояния постепенно утрачивают это свойство. Такое явление наглядно прослеживается на рассмотренном выше
примере с заменой березняка еловым лесом.
2. Биомасса органического вещества увеличивается по ходу сукцессии. Совершенно ясно, что с течением времени как в наземной, так и в водной среде, происходит накопление разлагающегося органического вещества. Естественные водоемы часто содержат в растворенном состоянии столько же органического вещества, сколько его находится в теле живых организмов.
3. Видовое разнообразие имеет тенденцию увеличиваться по ходу сукцессии. Это тесно связано со вторым аспектом, поскольку по мере роста биомассы растет число экологических ниш (лесная подстилка, гумус, кора, сухостой). В результате наиболее интенсивно увеличивается видовое разнообразие гетеротрофов. Автотрофы достигают этого на сравнительно ранних этапах сукцессии и обеспечивают максимум первичной продукции, создавая тем самым благоприятные условия для увеличения разнообразия гетеротрофов.
|
|
|
|
|
Дата добавления: 2014-11-06; Просмотров: 1639; Нарушение авторских прав?; Мы поможем в написании вашей работы!