КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
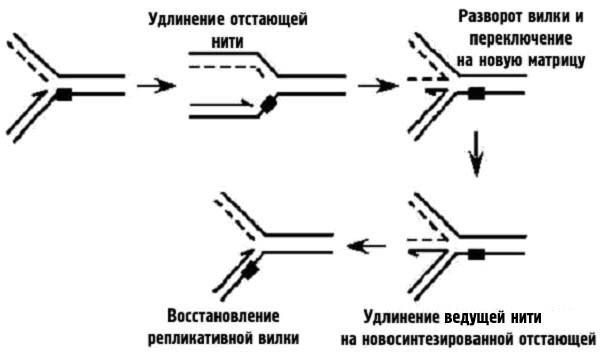
Модель прохода повреждения с переключением матрицы
|
|
|
|
Репарации поврежденных вилок репликации и ресинтез
Для репарации поврежденных вилок репликации существенна рекомбинация, которая снижает уровень мутагенеза, связанного с «перескакивающими» полимеразами. Доказано существование репликации, зависящей от рекомбинации в таких случаях, как конъюгация и трансдукция у бактерий, а у эукариот при мейозе, поддержании стабильности теломер и немутагенной репарации репликативных вилок. Документально подтверждено существование ресинтеза (рестарта репликации), связанного с рекомбинацией. В УФ-облученных клетках пекарских дрожжей в районе остановившихся вилок репликации образуются Холлидеевские структуры, а в выживших клетках возрастала частота генной конверсии и межгенной мититической рекомбинации. Одновременно было показано, что мутанты Rad6, Rad18 и Rad52 дефектны по пострепликативной репарации. Мутантный фенотип Rad6 и Rad52 при этом полностью соответствуют мутантному фенотипу Reca у E.coli. Вероятно, Rad52 играет ведущую роль в самом процессе рекомбинации, а Rad6 участвует в нем только на уровне регуляции. Нужно помнить, что для возникновения видимой Холлидеевской структуры требуется только отжиг цепей и ничего больше.
Гетеродимер Rad6/Rad18 совершенно необходим для работы любой системы, способной успешно завершать репликацию поврежденной ДНК. Мутанты по обоим этим генам крайне УФ-чувствительны и неповрежденные новосинтезированные нити ДНК в них после УФ-облучения не образуются.
Специфическая для структуры Холлидея геликазная-эндонуклеазная активность, способствующая ее разрешению, связана с комплексами Rad1/XPF и Mus81-Mms4. Одновременно оба эти комплекса способны к распознаванию остановившейся вилки репликации (выглядящей как вилка с «хвостом») и не проявляют сродства к обычным вилкам репликации. Вероятно, комплекс Mus81-Mms4 привлекается в район остановившейся вилки репликации белками чекпойнт-ответа, а его эндонуклеазная активность может участвовать в процессах, направленных на продолжение репликации. Для этого предложено множество моделей.
|
|
|
Белок Mgs1 (WHIP) является еще одним из возможных участников специального субпути PRR. Это ДНК-независимая АТФаза, способная образовывать комплекс с ДНК-полимеразой-δ и геликазой Sgs1 (так же как человеческий WHIP с WRN), появляется в районе остановившейся репликативной вилки и способствует проходу репликативным комплексом зоны повреждения.
Схематически эти модели изображены на рис. 39 и 40.
Первый способ, изображенный на рис. 39, основан на том, что при остановке вилки репликации перед повреждением на лидирующей нити, синтез на отстающей может продолжаться и приводить к появлению более длинного фрагмента вновь синтезируемой нити. Происходит разворот вилки с отжигом на сестринской хроматиде и продолжением синтеза лидирующей нити на вновь синтезированной отстающей. Как только лидирующая нить удлиниться так, чтобы перекрыть остановившее продвижение вилки повреждение, вилка разворачивается обратно и матричные нити спариваются со вновь синтезированными. Таким образом синтез продолжается, а повреждение оказывается обойденным.
На рис. 40 показано, что этот процесс может проходить и несколько иначе. При остановке вилки репликации перед повреждением на лидирующей нити, синтез на отстающей может продолжаться и приводить к появлению более длинного фрагмента вновь синтезируемой нити.
Эта вновь синтезированная нить может спариться со вновь синтезированной ДНК на лидирующей нити, так что вновь образованная ДНК на лидирующей нити сможет обойти повреждение. После прохождения поврежденного участка восстанавливается нормальная структура вилки репликации.
|
|
|
|
|
Дата добавления: 2014-11-06; Просмотров: 601; Нарушение авторских прав?; Мы поможем в написании вашей работы!