КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Обмен веществ и терморегуляция
|
|
|
|
6.10.1Факторы определяющие уровень обмена веществ в организме рыб;
Содержание кислорода в среде и интенсивность обмена. Одним из эндогенных факторов, влияющих на потребление кислорода, является мышечная активность. Согласно определению «основной обмен» – это потребление кислорода при относительном покое организма.
Таблица 17- Стандартный обмен
| Рыбы | Вес тела, г | Потребление кислорода, мл/(г*ч) | Температура окружающей среды, 0С |
| Илистый прыгун | 10-15 | 0,084(в морск. воде) 0,065(в пресн. воде) | |
| Серебряный карась | 0,042 | 200С | |
| Голец | 0,109 | 200С | |
| Чукучан Карликовый сомик Карп Солик Ictalurus | 43-127 | 0,109 0,092 0,067 0,07 | 150С 200С 200С 200С |
Таблица 18 - Потребление кислорода молодью серебряного карася в период активности (I) и в условиях стандартного обмена (II) при различных температурах.
| Вид рыбы | Потребление кислорода, мл/(г*ч) | ||
| Во время полета | В покое | При оцепенении | |
| Calyptc Selasphorus | 42,4 | 0,17 |
У серебряного крася эффективность использования кислорода Е, возрастает с увеличением скорости плавания V1,65 согласно уравнению;
Е=0,031 V1,65
На интенсивность обмена влияет не только мышечная, но и другая активность, например функция половой системы в период размножения. У рыб при возбуждении потребление О2 может значительно возрастать даже без заметного усиления мышечной активности.
У высокоактивных рыб, таких, как этеостома, интенсивность обмена в несколько раз больше, чем у малоподвижных видов. Это различие обнаруживается и на тканевом уровне: у активных рыб – потребление кислорода тканью головного мозга in vitro составляет от 11 до 14 мл/(кг.мин), у менее активных морского окуня и крокера от 7,7 до 9,7 и, наконец, у совсем малоподвижных – камбалы и опсануса –от 5,6 до 6,9 мл/(кг.мин.).
|
|
|
Температура и время года. У большинства пойкилотермных организмов интенсивность обмена растет или снижается вместе с изменением температуры тела, причем в физиологическом диапазоне температур составляет около 2,50С.
Интенсивность стандартного обмена у рыб постепенно повышается с температурой вплоть до летального уровня. Обмен в активном состоянии может либо повышаться с температурой до определенного уровня; а затем оставаться постоянным, либо проходить через максимальное значение при температуре, выше которой рыба уже не способна совершать интенсивную мышечную работу.
Разность между уровнями обмена в покое и в активном состоянии называется физиологическим резервом активности, которая возрастает с повышением температуры до некоторого максимума, а затем падает. Как резерв, так и сама активность достигают максимума приблизительно при одной и той же температуре. Водные пойкилотермные организмы, активные в различных диапозонах температур, обладают способностью к акклиматизации. Такая адаптация позволяет животному компенсировать сезонные изменения температуры или влияние географической широты.
Кумжа наиболее активна при 150С, кара – при 250С; для сомиков такой максимум не установлен. У высокоактивных костистых рыб хорошо развит механизм температурной компенсации.
Соленость. Пресноводные рыбы обычно потребляют больше кислорода, чем морские. Слегка солоноватой воде эвригалинные организмы (например, Myfilus edulis) потребляют больше кислорода, чем в морской воде, а стеностеногалинные (например, Asferias) меньше. Потребление кислорода жабрами в малосоленой воде возрастает.
Фотопериод и физиологические ритмы. У многих животных потребление кислорода подчинено суточному или лунному ритму. Фотопериод также может влиять на обмен. При 8-часовом фотопериоде Pachygrapsus потребляет на 55% больше кислород, чем при 16 часовом. При 9 часовом периоде у ушастого окуня выше, чем при 15 часовом, если температура воды не ниже 100С.
|
|
|
Гормоны. Обмен веществ очень часто зависит от пола животного, но эта зависимость не всегда одинакова. Влияние физических факторов на метаболизм во многих случаях опосредуется через гормоны. Экстракты щитовидной железы усиливают обмен у амфибии. Стероиды половых желез повышают интенсивность обмена у рыб.
Развитие. Стадия развития или жизненного цикла может сама по себе влиять на обмен, независимо от размеров тела. Как правило, с возрастом уровень обмена понижается, но на эмбриональной стадии интенсивность дыхания может быть невысокой.
Размеры тела и энергетические затраты. Потребление кислорода иногда выражают в расчете на одну особь, а чаще всего пересчитывают на 1 г. или 1 кг. веса тела (влажный или сухой вес), реже на единицу количества азота. Принято также выражать интенсивность дыхания в виде показательной функции от веса тела, что означает число миллилитров О2 (при стандартных давлении и температуре), потребляемых на 1 г сухого веса за 1 ч, а Vo2- количество О2 на 1 г сырого веса (или единицу объема ткани). Общее потребление кислорода растет с увеличением размеров животного, но при расчете на единицу веса оно снижается.
Таблица 19 - Зависимость интенсивность обмена от размеров тела.
| Животные (рыбы) | Размеры тела | Потребление О2 мл/(г*ч) |
| Лосось Oncorhuncus | 3-1400 | 0,97 (в акт.атм.) 0,78 (станд.обмен) |
| Серебряный карась | 100-400 | 0,85 |
| Серебряный карась | - | 0,86 |
| Cadus | - | 0,70 |
У некоторых животных кривая, отражающая зависимость интенсивности обмена от содержания кислорода в среде, становится более пологой при величинах Ро2 выше атмосферного уровня (т.е. выше 155 мм.рт.ст.). Эти животные занимают промежуточное положение между формами с зависимым и независимым типами дыхания. Характер зависимости обмена от Ро2 и положение критической точки Ркрит.. определяются рядом внешних и внутренних факторов, в том числе экологическими особенностями животного. При значениях Ро2 ниже Ркрит.
интенсивность обмена зависит от количества доступного кислорода; если даже Ро2 выше Ркрит., то лимитирующим фактором может быть концентрация субстратов, ферментов или кофакторов. Величины Ркрит. не во всех случаях постоянны, они могут несколько раз изменяться в зависимости от условий определения.
|
|
|
Зависимость обмена от давления О2, и механизмы этого влияния. Во время активности уровень обмена в большей степени зависит от Ро2, чем в период покоя. Максимальная скорость плавания, так же как и потребление кислорода, коррелирует с содержанием О2 в воде. У ушастого окуня величина Ркрит. растет с температурой, и при резком падении Ро2 она бывает выше, чем при медленном понижении. У рыбы Callionimus дыхание зависит от парциального давления О2: при снижении внешнего Ро2 частота дыхания и сокращении сердца уменьшается и потребление О2 падает. Рецепторы, чувствительные к кислороду, находятся в области жабр.
6.11 Теплообмен и терморегуляция Теплота - основа химических реакции, из которых складывается жизнедеятельность организма. Температурные условия оказываются одним из важнейших факторов, влияющих на интенсивность обменных процессов. Закономерность воздействия температуры на метаболизм выражается общим для всех химических реакций правилом Вант-Гоффа, согласно которому повышение температуры ведет к пропорциональному возрастанию скорости реакции.
В живом организме химические процессы всегда идут с участием сложных ферментных систем, в результате чего увеличивается их скорость и количественно меняется реакция на повышение температуры. Верхний температурный порог жизни теоретически определяется температурой свертывания белков. Необратимые нарушения структуры белков возникают при температуре порядка 600С, которую принято называть температурой «тепловой смерти».
Аналогичные нарушения метаболических и регуляторных процессов наступают и при очень низких температурах, нередко определяя собой нижний температурный предел жизни. Нарушения деятельности сердца при слабом охлаждении проявляются в ритме сокращений и сократимости сердечной мышцы, а при более сильном – в ее проводимости и возбудимости. При одном и том же снижении температуры удлинение периода диастолы выражено сильнее, чем систолы.
|
|
|
В результате различной реакции на температуру по достижении определенного порога (неодинакового у разных видов) возникает дискоординация функции в организме, ведущая к его гибели. При низкой температуре появляются структурные изменения в клетках и тканях, замерзание внеклеточной и внутриклеточной жидкости. По принципиальным особенностям теплообмена различают две крупные экологические группы животных: пойкилотермные и гомойотермные. К пойкилотермным (от греческого «poikilos» – «изменчивый», «меняющийся») относятся все животные, кроме птиц и млекопитающих.
Одно из наиболее заметных свойств пойкилотермных животных – неустойчивость температуры их тела, меняющийся в довольно широких пределах в зависимости от изменений температуры окружающей среды.
Температура тела. Характерная особенность теплообмена пойкилотермных животных заключается в том, что благодаря относительно низкому уровню метаболизма. Главным источником поступления тепловой энергии у них является внешнее тепло. Этим и объясняется зависимость температур тела пойкилотермных от температуры окружающей среды. Изменчивость температуры тела влечет за собой изменения скорости обменных реакции пойкилотермных животных.
Противоточный теплообмен. Теплообмен водных, наземных представлен противоточным теплообменом. В этом случае сосуды, по которым проходит холодная венозная кровь и возвращающаяся из плавников к телу, проходят рядом с артериями. Благодаря такому расположению артериальная кровь отдает тепло венозной крови, которая нагревается, прежде чем распределяться по телу. Кровь по артериям и венам течет в противоположных направлениях и между этими параллельными системами трубок происходит обмен тепла, получивший название противоточного теплообмена.
Подобная противоточная система существует между воздухом и кровью в жаберных лепестках рыб, благодаря чему артериальная кровь насыщается кислородом, сколько имеется ее в протекающей воде. Вода и кровь текут в противоположных направлениях. В результате этого рыбы извлекают из воды 80-90% содержащего в ней кислорода и создают условия для большего насыщения крови кислородом.
Энергетические затраты. Процесс метаболизма возрастает при активной мышечной работе у рыб в момент движения растраты 0,2 на 1 гр (кг) веса составит около 78 мл/км.
Энергетические затраты при плавании очень низкие вследствие того, что удельный вес тела близок к удельному весу воды, а энергия затрачивается только на преодоление сопротивления воды. При повышении жидкости в организме повышается сопротивляемость воды, что приводит к увеличению затрат энергии. У мелких рыб затраты энергии больше чем у крупных. Благодаря своей форме и площади тела, крупные рыбы легко передвигаются с меньшими усилиями.
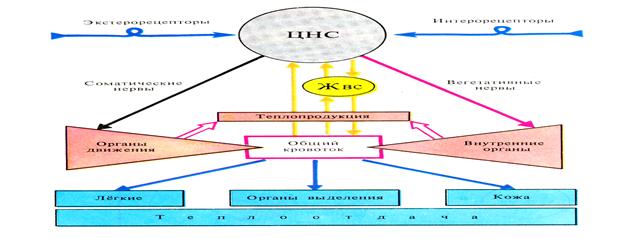
Рис. 50 Схема регуляции температуры тела у теплокровных животных (по Кулланде,1970).
В момент активного плавания в мышцах тунцов и акул создается высокая температура, хотя температура тела у рыб очень близка к температуре окружающей воды. Кровь, через жабры обогащаясь кислородом, охлаждается до температуры воды, вследствие того, что артериальная кровь от жабр течет по сосудам лежащим близко к поверхности тела рыбы. В толщах мышц, где температура тела, может быть на 140С выше температуры воды, находятся сосуды, поддерживающие внутреннюю температуру.
В процессе обмена веществ непрерывно происходит изменение энергии. Энергия сложных химических соединений (потенциальная) при расщеплении освобождается и превращается в тепловую энергию. Часть энергии в организме превращается в механическую и незначительное количество в электрическую.
Контрольные вопросы.
1.Дайте определенике понятий «аноболизм» и «катоболизм», их этапы,
единство и различия.
2.Охарактеризуйте роль желудочно-кишечного тракта,печени, почек и мышц
в обмене веществ.
3.Что такое «азотистый баланс»?
4.Перечислите особенности и основные этапы углеводного, жирового,
белкового, солевого, минерального, водного обмена у рыб.
5.Охарактеризуйте механизм превращения энергии в организме рыб
6.Что такое прямая и непрямая колориметрия?
7.Что такое основной обмен, как он определяется?
8.Какие факторы влияют на интенсивность теплопродукции и теплоотдачи?
9.Механизм регуляции обменных процессов.
ГЛАВА 7
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 1337; Нарушение авторских прав?; Мы поможем в написании вашей работы!