КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Краткая характеристика ионных токов сердца
|
|
|
|
Na+ ток - основной ток в сердечной мышце. Самый значительный ионный ток- INa, - это поток ионов через натриевые каналы, поскольку их плотность составляет 200 Na+-каналов на квадратный микрометр мембраны. Они есть в мышцах предсердий и желудочков, в волокнах Пуркинье. В кардиомиоцитах синоатриального и атриовентрикулярного узлов этот ток отсутствует.
INa обусловлен классическими потенциал-зависимыми Na+-каналами, имеющими a- и b1-субъединицы. a-субъединица Na+-каналов сердечной мышцы имеет места для фосфорилирования, что делает ее чувствительной к стимуляции через цАМФ-зависимую протеинкиназу. В покое (при отрицательном МП) эти каналы закрыты, однако они способны мгновенно активироваться (в течение 0,1 - 0,2 мс) в ответ на локальную деполяризацию, вызываемую распространяющимся ПД, и создают массивный входящий ток. Этот ток лежит в основе восходящей фазы ПД (рис. 2.6Б, фаза деполяризации). Когда МП становится положительным, эти каналы закрываются, то есть идет процесс инактивации, который развивается медленнее, чем процесс активации, но все же достаточно быстро. Во время фазы плато ПД, когда МП поддерживается на уровне, немного выше 0 (фаза 2), этот ток не исчезает полностью, и дает небольшой, но важный вклад в поддержание фазы “плато”.
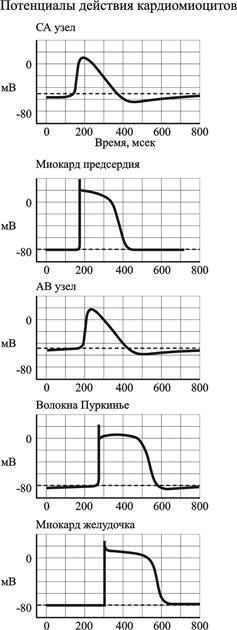
Рис. 2.7. Потенциалы действия различных участков сердца
Са2+ ток кардиомиоцитов обеспечивается преимущественно каналами L-типа. Кальциевый ток присутствует во всех кардиомиоцитах. Са 2+-каналы L-типа являются доминирующими в мембране сердечных клеток, хотя обнаружены еще и каналы Т-типа, однако количество их невелико. В синоатриальном узле ICa участвует в развитии пейсмекерной активности. В синоатриальном и в атриовентрикулярном узлах ICa является входящим током, который обусловливает восходящую фазу (фаза деполяризации) ПД. А поскольку эти клетки лишены “быстрых” Na+-каналов, то восходящая фаза развивается медленнее, чем в мышечных клетках предсердий и желудочков (сравните рис. 2.6 А и Б). Следовательно, крутизна нарастания спонтанной диастолической деполяризации мембраны происходит здесь с меньшей скоростью, что приводит к так называемой “ электрической задержке ” проведения возбуждения в атриовентрикулярном узле и задержке сокращения желудочков относительно сокращения предсердий. Эта особенность обеспечивает необходимое время для того, чтобы сначала сократились оба предсердия и эффективно протолкнули кровь в желудочки, и только после этого наступило сокращение обоих желудочков.
|
|
|
ICa , хотя и в гораздо меньшем объеме, участвует также (вместе с INa) и в развитии восходящей фазы быстрой деполяризации ПД мембраны рабочих кардиомиоцитов предсердий и желудочков, а также атипических кардиомиоцитов волокон Пуркинье, что способствует более быстрому проведению ПД в этих тканях. Са2+-каналы, как и Na+, закрыты при отрицательном МП и при более положительных значениях быстро активируются (в течение 1 мс) и затем инактивируются (время полуспада – 10 - 20 мс). Са2+ ток присутствует и во время фазы 2 ПД, где он участвует в формировании фазы плато. В рабочих кардиомиоцитах миокарда предсердий и желудочков ионы Са2+, входящие через L-тип каналов, активируют освобождение Са2+ из саркоплазматического ретикулума по типу Са-зависимого освобождения Са2+. Блокаторы L-типа Са2+ каналов - верапамил, дилтиазем и нифедипин ингибируют ICa, и используются как терапевтические препараты.
Реполяризующий К+ ток развивается медленно. Длительность ПД сердечной мышцы на два порядка (примерно в 100 раз) больше, чем длительность ПД скелетных мышц. Поэтому, реполяризующий К+ ток - IК, развивается медленнее, со значительной задержкой; он обнаружен во всех кардиомиоцитах, и отвечает за реполяризацию мембраны в конце ПД (фаза 3 рис. 2.6 А и Б). При отрицательных значениях МП поток ионов К+ невелик, при деполяризации К+ каналы активируются медленно - в течение 20 - 100 мс, но не инактивируются, в отличие от других каналов. В клетках синоатриального и атриовентрикулярного узлов этот ионный ток участвует в развитии спонтанной диастолической деполяризации.
|
|
|
Неселективный катионный ток - If. Так называемый “пейсмекерный ток” - If , обнаружен в мембранах кардиомиоцитов синоатриального и атриовентрикулярного узлов и в волокнах Пуркинье (рис. 2.6 А). Обусловлен этот ток работой неспецифического катионного H-канала (hyperpolarization-activated - то есть активируемого гиперполяризацией). Поскольку через этот канал идут и К и Na ионы, потенциал реверсии для If каналов равен -20 мВ, то есть находится между потенциалами равновесия для К+ (-90 мВ) и для Na+ (+50 мВ). Н-каналы имеют необычные свойства (поэтому их обозначили буквой “f” от слова “funny” - смешной, забавный), они активируются при гиперполяризации мембраны в конце фазы 3 ПД. Активируются они медленно (время активации - 100 мс) и не инактивируются, Н-каналы обеспечивают входящий, деполяризующий ток по мере того, как они медленно активируются в конце фазы 3. Однако If не единственный ток, отвечающий за пейсмекерную активность, в развитие фазы 4 в синоатриальном и атриовентрикулярном узлах вносят вклад также ICa и IK.
Рабочие кардиомиоциты предсердий и желудочков. Необходимо отметить, что МП таких кардиомиоцитов значительно более отрицателен (-80 -90 мВ), чем МП атипических кардиомиоцитов проводящей системы (рис. 2.7.). Следовательно, возбудимость не обладающих пейсмекерной активностью кардиомиоцитов гораздо ниже, чем у пейсмекерных клеток.
Кардиомиоциты предсердий. В пределах предсердий ПД распространяется непосредственно от клетки к клетке. ПД сократительных или рабочих кардиомиоцитов - это результат сложения трех потенциалозависимых мембранных токов: 1) INa 2) IK и 3) ICa. Они не обладают в норме спонтанной (то есть пейсмекерной) активностью. Предсердия снабжены 4-мя специальными проводящими пучками (рис. 2.8.). Один из них - пучок Бахмана (передний межпредсердный миокардиальный пучок), проводит импульсы от синоатриального узла к левому предсердию. Три других интернодальных (межузловых) пучка - передний, средний и задний проводят ПД от синоатриального к атриовентрикулярному узлу. Следовательно, первый шаг в распространении ПД - это деполяризация предсердий в направлении, совпадающем с анатомической осью сердца, - справа налево и сверху вниз.
|
|
|
Кардиомиоциты желудочков. Последовательность событий при активации желудочков занимает в общей сложности 100 мс. Единственный путь электрической связи предсердий и желудочков проходит через атриовентрикулярный узел.После того, как ПД достигнет атриовентрикулярного узла, он распространяется по пучку Гиса, волокнам Пуркинье и входит в миокард желудочков, где ПД передается от клетки к клетке. Следующий этап- деполяризация межжелудочковой перегородки слева направо, затем развивается деполяризация переднего отдела перегородки. Миокард всегда деполяризуется в направлении от эндокарда (клетки, выстилающие желудочки) к перикарду (клетки наружной поверхности сердца). Миокард левого желудочка деполяризуется в направлении к верхушке сердца, в то время как волокна Пуркинье проводят ПД к основанию левого желудочка.

Рис. 2.8. Проводящая система сердца.
Затем волна деполяризации распространяется от верхушки к основанию желудочков по волокнам Пуркинье. Последней деполяризуется задняя поверхность левого желудочка.
В кардиомиоцитах желудочков обнаружены 3 основных потенциал-зависимых ионных тока: 1) INa 2) IK и 3) ICa (рис. 2.6 Б). Здесь нет тока If, следовательно, в норме эти кардиомиоциты не обладают пейсмекерной активностью. Начинаясь от уровня МП в -80 мВ, восходящая фаза быстрой деполяризации в миокарде желудочков является следствием активации INa каким-то внешним стимулом (например, импульсом, проводимым по волокнам Пуркинье или пришедшим из соседней клетки). Особенно значимым для мышцы желудочков является Са2+ ток, т.к. он обеспечивает вход ионов Са2+, активирующих освобождение Са2+ из саркоплазматического ретикулума. Фазы быстрой реполяризации (фаза 1), плато (фаза 2) и реполяризации (фаза 3) обусловлены теми же механизмами, что и в миокарде предсердий (рис. 2.6. Б).
|
|
|
2.1.2. Рефрактерный период сердечной мышцы
Сердечная мышца, как и все электровозбудимые ткани, во время развития потенциала действия нечувствительна к новому возбуждению, то есть рефрактерна. Следовательно, рефрактерный период сердца – это интервал времени, в течение которого нормальный сердечный импульс не может повторно возбуждать уже возбужденную область сердечной мышцы. Это период абсолютной рефрактерности. В норме период абсолютной рефрактерности желудочков длится от 0,25 до 0,3 с, что соответствует продолжительности потенциала действия (0,3 с или 300 мс). Затем наступает период относительной рефрактерности, длительностью примерно 0,05 с, в течение которого мышца уже восстанавливает свою возбудимость, но она остается все еще пониженной. В этот период мышца может быть возбуждена, но только в ответ на очень сильный стимул. В самом конце периода рефрактерности наблюдается короткий период гипервозбудимости (так называемый “ уязвимый” период). Рефрактерный период мышцы предсердия намного короче и составляет примерно 0,15 с, а период относительной рефрактерности – 0,03 с. Поэтому предсердия могут сокращаться с большей частотой, чем желудочки. Длительность ПД кардиомиоцитов, а следовательно и период рефрактерности, не является строго фиксированной константой и может укорачиваться при увеличении частоты сердечных сокращений.
Ионные механизмы рефрактерности. Как уже говорилось, однократно активированные кардиомиоциты желудочков будут рефрактерны к повторной активации. Этот период абсолютной рефрактерности объясняется инактивацией входящих токов - INa и ICa, поэтому дополнительный электрический стимул не будет иметь никакого эффекта. В конце фазы плато клетка начинает реполяризоваться, т.к. начинает увеличиваться IK. По мере восстановления ICa и INa после инактивации наступает период относительной рефрактерности. В этот период дополнительный электрический стимул может вызвать генерацию ПД, но гораздо меньшей амплитуды.
Рефрактерность защищает сердце от возможного сокращения в ответ на частые повторные стимулы, которые могут приходить, например, из эктопических (расположенных вне проводящей системы) очагов генерации ПД. Рефрактерность также предотвращает развитие в сердечной мышце тетанусов (сильное, длительное сокращение), наблюдаемых в скелетной мускулатуре. Экстрасистолические сокращения снижают эффективность насосной функции сердца, а тетанусы могли бы привести к тому, что сердце все время находилось бы в систоле.
|
|
|
|
|
Дата добавления: 2014-11-08; Просмотров: 1521; Нарушение авторских прав?; Мы поможем в написании вашей работы!