КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Локализация функций
|
|
|
|
Первоначальные наблюдения Бергера (Berger, 1929) базировались на убеждении, что ЭЭГ отражает общую активность мозга. Психофизиологи в общем склонны были разделять это мнение. Отсюда следовало, что, если электроды расположены в области черепа, точное положение их не столь существенно. А это в свою очередь подразумевает «холистическую» точку зрения на функции мозга в противоположность представлению о специфической локализации функций.
Когда Галль выдвинул свою систему френологии, она оказалась, как сказал Эдмунд Дж. Боринг в своей «Истории экспериментальной психологии» (Boring, 1950), «примером теории, которая, будучи в основном ошибочной, была верной ровно настолько, чтобы способствовать прогрессу научной мысли». Ибо Галль не только пробудил интерес к анатомии коры мозга, но воскресил также идею о локализации функций и распространил ее на сложные психические процессы.
За полтора столетия, прошедшие с тех пор, представление о локализации функций несколько раз входило в фавор и впадало в немилость. Исследования Пьера Флуранса в начале XIX века говорили в пользу холистического взгляда. На основе наблюдений, которые оказались ошибочными, он сделал вывод о диффузном представительстве сложных психических функций в мозгу. Эта концепция «эквипотенциальности» различных участков мозга властвовала в западной медицине вплоть до 1861 года, когда Поль Брока на заседании Парижского антропологического общества представил результаты своих наблюдений над больными с повреждениями мозга. Он продемонстрировал препараты мозга нескольких больных, которые после апоплексии потеряли способность говорить и у которых были ограниченные повреждения в левом полушарии. Сходные клинические данные Вернике и других исследователей воскресили идею локализации функций в мозгу. Примерно в тот же период теория локализации получила подкрепление в экспериментах Фрича и Гитцига, показавших, что электрическое раздражение определенных участков коры
|
|
|
Глава 9
Головной мозг
вызывало у собаки совершенно определенные мышечные движения.
В 1905 году Пьер Мари повторно исследовал препараты, представленные Брока; и показал, что его выводы тоже были основаны на ошибочных наблюдениях — что поражения мозга были значительно более обширными, чем думал Брока. Снова поддержку получила холистическая точка зрения. Обширные исследования Карла Лэшли, который в первой половине этого столетия изучал влияние повреждений мозга на результаты научения крыс в лабиринте, также подкрепляли холистическую точку зрения. Данные Лэшли указывали на то, что ухудшение функций зависит от общего объема разрушенной мозговой ткани, а не от повреждения специфических участков. Лэшли абсолютизировал представление об эквипотенциальности — любая часть коры, утверждал он, может принять на себя функцию любой другой. Однако даже он постепенно пришел к признанию того, что в коре есть некоторое «разделение труда».
, Сегодня вопрос состоит не в том, существует ли локализация функций в мозгу человека, а в том, насколько сильно она выражена. Один из наиболее авторитетных современных исследователей в этой области — советский нейрофизиолог А. Р. Лурия (1973) —выдвинул представление о том, что мозг организован как система функциональных единиц, координирующих сложные психические процессы. Во внутренней организации каждой единицы может существовать значительная пластичность, однако сами единицы связаны с определенными участками коры. Другие исследователи продолжают придерживаться мнения о большей или же о меньшей степени локализации.
|
|
|
Все, что мы знаем о поведении человека, говорит нам, что мозг — это не аморфное скопление клеток. Он должен обладать упорядоченной организацией, чтобы была возможна вся сложная деятельность человека. В этом отношении мы только начинаем понимать язык мозга. По-видимому, внутримозговая специализация связана не просто с каким-то анатомическим распределением клеток, а скорее со специфическими паттернами их разрядов'(John, 1972; Bartlett, John, 1973). Через сто лет наши нынешние представления о локализации функций наверняка будут казаться такими же грубыми, какими представляются нам сегодня френологические карты Галл я.
Именно в этой области уникальный вклад могут внести исследования ЭЭГ. Каким бы примитивным ни был в настоящее время метод регистрации электрической активности, она отражает функционирование мозга здорового человека. Фактически все наши современные представления о локализации
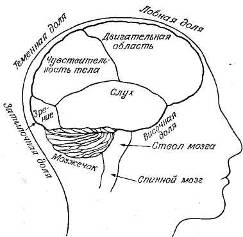
Рис. 9.2. Главные отделы мозга.
основаны на изучении животных и больных с поражениями мозга. Вводные сведения по этому вопросу превосходно изложены у Гарднера (Gardner, 1974) и Роуза (Rose, 1976). Изучение локальных ЭЭГ может расширить и дополнить имеющиеся данные.
Анатомически кору обычно подразделяют на четыре главных отдела, или доли, как это показано на рис. 9.2. Во всяком случае, ясно, что разные сенсорные модальности представлены в этих долях по-разному.
Затылочная кора участвует в обработке зрительных стимулов. Именно от нее психофизиологи чаще' всего отводили ЭЭГ, так как альфа-ритм обычно имеет здесь наибольшую амплитуду. Височная кора больше связана со слуховой системой. Поэтому, если нас интересует реакция ЭЭГ на серию тонов, то лучше будет, видимо, использовать височное, а не затылочное отведение. Соматосенсорная кора занимает небольшую область вокруг центральной борозды — углубления, разделяющего лобную и теменную доли. Именно сюда поступает осязательная информация от всего тела и отсюда посылаются двигательные команды к мышцам.
Левая и правая стороны мозга несколько различаются. Хотя многое дублируется в обеих половинах мозга, в целом наша нервная система основана на принципе перекрестной иннервации: левое полушарие мозга управляет правой половиной тела, а правое — левой. Поэтому можно ожидать, что при сжимании правого кулака изменения ЭЭГ будут более заметными в левом полушарии, и наоборот. Так оно и есть на самом
|
|
|
Глава 9
Головной мозг
деле. Наконец, между функциями левого и правого полушарий существуют еще различия в психологических функциях. У правшей, например, лингвистические функции, как и предполагал Брока, связаны с левым полушарием. Мы вернемся к этому позже, когда будем рассматривать новейшие данные о межполушарной асимметрии.
Существует много данных в пользу того, что изменения ЭЭГ, регистрируемые с различных участков головы, отражают активность соответствующих областей мозга. Например, Гасто (Gastaut, 1952) изучал в сенсомоторной области коры то, что теперь называют сенсомоторным ритмом. Этот ритм по форме похож на альфа-ритм затылочной области. Однако сенсомоторный ритм блокируется, когда человек шевелит конечностями, в то время как затылочный альфа-ритм в большей степени реагирует на зрительное внимание. Таким образом, хотя мы еще не знаем в точности механизм генерирования ритмов ЭЭГ на клеточном уровне, мы знаем, что ритмы, отводимые от разных участков черепа, обладают различной реактивностью. Есть надежда, что исследование локальных ЭЭГ поможет нам в понимании организации мозга. А изучение природы такого «биологического разделения труда» в свою очередь поможет психологам разработать биологически обоснованную классификацию форм поведения.
Расположение электродов
Мы знаем, что активность, одновременно регистрируемая с разных точек головы, может сильно различаться. Если мы поставим этот простой факт в связь с проблемой локализации, то увидим, насколько важно для электроэнцефалографии понимание основных принципов расположения электродов.
Как при всякой регистрации электрической активности, для образования замкнутой цепи следует помещать на теле два электрода. При записи ЭЭГ используются два основных метода. При биполярной записи оба электрода располагают на скальпе. В таком случае каждый из них будет улавливать электрическую активность мозга, а полученная ЭЭГ будет отражать разность потенциалов между двумя электрическими сигналами. При монополярной записи один электрод помещают на скальпе, а другой — на «электрически нейтральной» точке (чаще всего на одной из ушных мочек). На первый взгляд предпочтительнее кажется монополярная регистрация, так как можно было бы думать, что при этом отводится активность трлько одной точки мозга. Однако есть основания полагать, что биполярная запись дает нам информацию о более локальных явлениях (Lindsley, Wicke, 1974). В настоящее время относи-
|
|
|
тельные достоинства моно- и биполярной записи все еще остаются предметом дискуссии.
Тем не менее существует общее соглашение относительно системы точного расположения электродов. В ранних работах положение электродов часто указывали, просто называя соответствующую долю мозга, например «правая затылочная» Чтобы избежать расхождений между разными лабораториями, Международная федерация обществ электроэнцефалографии и клинической нейрофизиологии приняла систему «10—20», позволяющую точно указывать расположение электродов. Эта система представлена на рис. 9.3.
В соответствии с этой системой у каждого испытуемого точно измеряют расстояние между назионом (вдавление на переносице) и инионом (твердый костный бугорок на затылке), а также между правой и левой ушными ямками. Возможные точки расположения электродов разделены интервалами, составляющими 10 или 20% этих расстояний на черепе. Дополнительные пояснения даны в приложении Е. Для того чтобы нам можно было указывать положение электродов в разных исследованиях, отметим здесь еще несколько моментов. Буквы перед цифрами означают соответствующую долю мозга: О — затылочная доля, F — лобная, Р — теменная, Т — височная, С — область центральной борозды. Нечетные номера мест отведения относятся к левому полушарию, четные — к правому. Буквой Z обозначается отведение от верхушки черепа. Это место, которое часто используют, называется также вертексом.
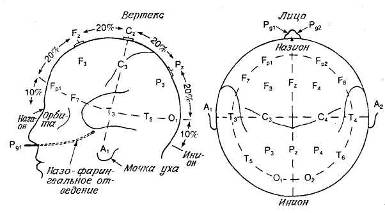
Рис. 9.3. Система 10—20. (Jasper, 1958.)
Расположение электродов на поверхности головы. F — лобная область. С — центральная, Р — теменная, Т — височная, О — затылочная. Нечетные индексы — левая половина головы, четные индексы — правая, Z — средняя линия.
144 Глава 9
Головной мозг
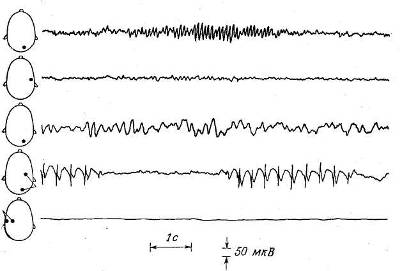
Рис. 9.4. Типичные электроэнцефалограммы. (Vander G., Kempe L., A Primer of Electroencephalography, Roche Laboratories, Division of Hoffman — La Roche, Inc., 1970.)
На схематических изображениях головы слева от каждой записи показано расположение электродов. Две первые записи сделаны одновременно у здорового испытуемого в состоянии покоя. Обратите внимание на высокоамплитудные альфа-волны, регистрируемые только в затылочном отведении (вверху). Третья запись —от затылочной области у здорового ребенка. У маленьких детей альфа-ритм отсутствует. На следующей записи видны необычные пики, связанные с малым эпилептическим приступом. Нижняя запись демонстрирует отсутствие электрической активности при «мозговой смерти» (хотя сердце этого больного продолжало биться, клинически он был мертв).
Применение такой системы напоминает нам, что мы не регистрируем, как это думал Бергер, «ЭКГ головы». Разные отделы мозга реагируют по-разному.
Анализ ЭЭГ
Регистрируя ЭЭГ во время сна, можно за одну ночь исписать мозговыми зигзагами больше полумили ленты полиграфа. Как свести эту гору данных к приемлемому объему?
Мы уже вкратце говорили об основных ритмах ЭЭГ (дельта, тета, альфа и бета) и о принципах частотного и амплитудного анализа. Вернемся несколько назад, чтобы рассмотреть клиническую интерпретацию ЭЭГ.
На рис.,9.4 показаны примеры нормальной и патологической ЭЭГ. В клинике, вероятно, наибольшее значение имеет
применение электроэнцефалографии для диагностики малых эпилептических припадков (petit mal). Это состояние трудно определить по поведению, поскольку его признаки — кратковременная потеря внимания, спутанность речи и т. п.— могут быть относительно слабо выражены. В таких случаях ЭЭГ особенно полезна для оценки эффективности приема больным тех или иных противосудорожных препаратов.
При записи ЭЭГ в клинике применяют достаточно стандартное расположение электродов и стандартный набор заданий, в число которых входят гипервентиляция и навязывание светового ритма, когда с помощью ритмических вспышек света искусственно вызывают судорожную активность. Поскольку при эпилепсии наблюдаются резкие изменения ЭЭГ, у клинических электроэнцефалографистов не возникало необходимости в точных критериях нормы и патологии.
Для некоторых исследователей чтение ЭЭГ стало скорее искусством, чем наукой. Воодушевленные большими успехами в выявлении аномально функционирующих участков мозга у эпилептиков, эти исследователи перешли к попыткам найти физиологические отклонения у людей с психическими аномалиями. При этом отсутствие точности в физиологическом подходе совмещалось у них с такой же неточностью при описании изменений поведения, когда были начаты исследования электроэнцефалографических коррелятов различных психозов, алкоголизма, преступности и т. п. Однако алкоголизм, психозы, преступность определялись в разных работах по-разному, а «физиологические корреляты» часто бывали просто субъективными оценками «аномальности». Не удивительно поэтому, что, несмотря на сотни таких экспериментов, не удалось прийти к какому-либо общему мнению относительно связи между психическими отклонениями и «необработанной» ЭЭГ.
В связи с этим многие исследователи ощущали необходимость в более точном подходе, чем простое рассматривание ЭЭГ. Было предложено много методов и еще больше постоянно разрабатывается. Краткое описание наиболее популярных методов дает Шагасс (Shagass, 1972), а мы ограничимся здесь лишь общим обзором возможных подходов. Сначала мы должны провести различие между методами, анализирующими одиночные и множественные отведения.
В случае одиночных отведений нас интересует изолированное рассмотрение записей, сделанных от разных отделов мозга (отведения могут быть моно- и биполярными). Для разграничения двух подходов существенно то, что в данном случае мы рассматриваем каждый раз только один канал информации. В более сложном случае — при множественных отведениях —
Глава 9
мы имеем дело с фазовыми отношениями между двумя или несколькими отделами мозга, поэтому здесь необходима одновременная регистрация ЭЭГ на нескольких каналах.
И опять-таки наиболее простая форма анализа отдельных отведений — это обычный клинический метод визуального рассмотрения записей в поисках хорошо заметных особенностей. Можно достигнуть большей точности, если анализировать активность в разных частотных диапазонах. Мы можем, например, осуществлять фильтрацию ЭЭГ таким образом, что на втором канале полиграфа будет регистрироваться только активность с частотой 8—13 Гц (альфа-ритм). Как уже говорилось, мы можем считать""какую-то минимальную ее амплитуду за «наличие альфа-ритма» и с помощью электроники или вручную определять процент времени, когда альфа-ритм (или другая полоса частот) превышает эту величину.
Более точные методы позволяют учитывать амплитуду активности в широком спектре частотных полос. Такие методы чаще всего основаны на математическом принципе рядов Фурье. Примерно 200 лет назад Фурье показал, что волна любой сложной формы математически идентична сумме синусоидальных волн разной амплитуды и частоты.
Таким образом, сложнейшие по форме волны ЭЭГ можно с помощью преобразования Фурье свести к ряду синусоидальных волн с разными амплитудами и частотами. Анализ спектра мощности дает картину относительной мощности разных частот (которая зависит от амплитуд синусоидальных составляющих). Математическое описание этих процессов очень сложно и требует применения быстродействующих цифровых вычислительных машин. Для нас сейчас важно то, что такой анализ дает значительно более полное описание ЭЭГ, нежели простой подсчет времени, которое занимает активность в том или ином частотном диапазоне. По существу, мы получаем итоговую картину распределения активности на широком диапазоне частот.
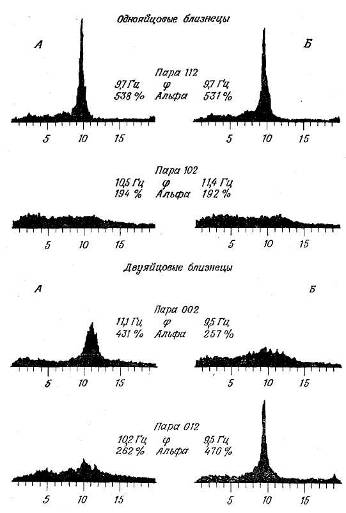
Этот метод хорошо иллюстрируют генетические исследования Ликкена и сотр. (Lykken et al., 1974) относительно ЭЭГ покоя. Производилось сравнение спектров мощности ЭЭГ у пар однояйцовых (идентичных) и двуяйцовых (неидентичных) близнецов. (Как и во многих других исследованиях по генетике человека, исходным предположением было то, что если генетически идентичные однояйцовые близнецы обнаруживают большее сходство по данному признаку, чем двуяйцо-вые, то этот признак считается наследственным.) Основной целью работы было изучение восприимчивости близнецов к гипнозу; при этом в начале и в конце одночасового эксперимента испытуемых просили сидеть три минуты с закрытыми
| Головной Most |
|
Рис. 9.5. Спектр мощности ЭЭГ у моно- и дизиготных близнецов в покое. (Lykken et al., 1974.)
Обратите внимание на то, что у однояйцовых близнецов записи очень сходны, а у двуяйцовых различаются. Это говорит о том, что параметры ЭЭГ покоя находятся под сильным влиянием наследственности. Ф — средняя частота альфа-полосы; уровень альфа-ритма (величина пика <?) выражен в процентах к мощности на частоте 3 Гц.
глазами. Как видно из рис. 9.5, между однояйцовыми близнецами обнаруживается разительное сходство. В самом деле, кривые близнецов сходны друг с другом почти так же, как сходны между собой записи каждого из них в начале и в конце
I
Глава 9
Головной мозг
опыта. Между тем в ЭЭГ двуяйцовых близнецов сходства не обнаруживается. Коэффициент наследуемости шести различных параметров ЭЭГ оказался равным в среднем 0,82 (определенный в той же работе коэффициент наследуемости сердечного ритма в покое составил 0,67).
Однако вернемся к обзору методов анализа. Использование вычислительных машин позволяет осуществить еще ряд подходов к анализу относительного уровня активности в разных диапазонах частот. Из них чаще всего применяются периодический анализ (или «метод пересечения нуля») и метод автокорреляции. Однако наиболее популярными методами изучения ЭЭГ остаются применение фильтров для разных частот и анализ спектра мощностей.
При анализе множественных отведений применяются гораздо более сложные приемы. Это порождает некоторые надежды, так как мы знаем, что «трескотня» нескольких миллиардов нервных клеток, координирующих наши психические процессы,— это сложное явление, требующее комплексного подхода. Если бы ЭЭГ состояла из ряда прямых линий, ее, возможно, было бы намного легче интерпретировать, но тогда она не обещала бы нам помощи в нашем стремлении приоткрыть окно в нашу внутреннюю жизнь.
Одним из самых перспективных методов анализа множественных отведений является метод, разработанный Ливановым и его сотрудниками (1966). Записывая ЭЭГ одновременно по 50 каналам, они вычислили 1225 коэффициентов корреляции (между всеми возможными парами электродов), чтобы установить, какие области мозга работают согласованно, т. е. в каких именно участках ЭЭГ относительно синхронна. Приведем один из важнейших результатов: у здорового человека в состоянии покоя ЭЭГ большинства областей коры не синхронна, тогда'как во время решения в уме арифметической задачи активность многих корковых областей, особенно лобной коры, синхронизируется.
Технические сложности при проведении одновременной записи по 50 каналам и анализе всех возможных корреляций должны быть поистине колоссальными. Однако весьма вероятно, что для успеха в понимании скрытых функций человеческого мозга потребуется именно такого рода смелость, решимость охватить всю эту сложность, вместо того чтобы отступить перед ней.
Сон
У здорового человека только циклические изменения сна легко выдали электроэнцефалографисту тайну своего суще-
ствования. Около трети жизни (т. е. 23 полных года, если мы проживем до 69 лет) мы проводим во сне. Однако эта столь ' значительная часть нашей жизни привлекла серьезное внимание ученых лишь сравнительно недавно — в результате прогресса электроэнцефалографии (и регистрации движений глаз, как уже говорилось в гл. 7).
Описывая сон с быстрыми движениями глаз (БДГ), мы упоминали о том, что существует несколько типов сна. Там, где случайный наблюдатель в течение восьми часов будет видеть только неподвижное тело, электрофизиолог сможет построить график переходов из одной фазы сна в другую. Еще в 1862 году Кольшюттер показал, что громкость звука, способного разбудить спящего человека, систематически изменяется в течение ночи. Но для того чтобы открыть четыре стадии сна, нужно было дождаться эры электроэнцефалографии.
На рис. 9.6 показана типичная запись затылочной ЭЭГ в четырех стадиях сна, описанных Дементом и Клейтманом (Dement, Kleitman, 1957).
Засыпая, человек обычно последовательно проходит через эти стадии. Любая помеха, например неожиданный шум, может повернуть процесс вспять, возвратить спящего назад на более раннюю стадию. В течение ночного сна человек совершает переходы между этими стадиями туда и обратно,
Стадия 1
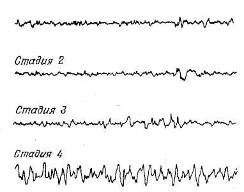
Рис. 9.6. Электрофалографические стадии сна.
В стадии 1, наступающей сразу, как только человек переходит ко сну, альфа-активность постепенно сменяется низковольтными изменениями потенциала. «Сонные веретена» — нерегулярные вспышки активности частотой 12—14 Гц — впервые появляются в стадии 2. Эти веретена вместе с высоковольтными дельта-волнами, достигающими 300 мкВ, наблюдаются и в стадии 3. И наконец, в стадии 4 в ЭЭГ преобладают высоковольтные дельта-волны.
Глава 9
Головной мозг
периодически погружаясь в стадию БДГ со сновидениями, которая по типу ЭЭГ очень сходна с 1-й стадией. У каждого человека есть свои особенности в смене этих стадий.
Есть некоторые указания на то, что эти стадии сна функционально различны. Например, Люс и Сегал (Luce, Segal, 1966) высказывают предположение, что 4-я стадия, наиболее характерная для первых часов сна, выполняет функцию восстановления физических сил. Если вы целый день грузили ящики, то вы скорее всего проведете в 4-й стадии больше времени, чем если вы весь день читали «Введение в психофизиологию». Внешние различия между физиологически столь разными периодами сна продолжают привлекать внимание исследователей (Snyder, Scott, 1972).
Изменения ЭЭГ были обнаружены и при лишении сна. Если вы всю ночь просидели за подготовкой к экзамену, у вас, вероятно, будут короткие периоды отключения внимания («микросон»). В те несколько секунд, на которые вы отключаетесь, ЭЭГ принимает вид, характерный для спящего человека (Liberson, 1945). В этом смысле ЭЭГ оказывается индексом «умственной активации» человека.
Именно такая отчетливая смена фаз в ЭЭГ во время сна пробудила в начале 50-х годов новый интерес к «теориям активации» применительно к поведению человека (см., например, Lindsley, 1951). Напомним, однако, что на протяжении всей книги при рассмотрении физиологии человека мы постоянно подчеркивали, как важно понимать особенности каждой из систем организма. Именно поэтому концепция активации представляется нам слишком упрощенной.
|
|
|
|
|
Дата добавления: 2014-11-16; Просмотров: 613; Нарушение авторских прав?; Мы поможем в написании вашей работы!