КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
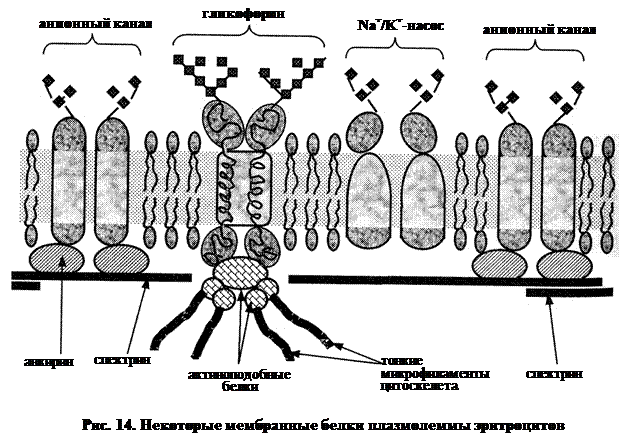
Некоторые белки плазмолеммы эритроцита
 В качестве наиболее ярких примеров структурных и транспортных белков ниже будут рассмотрены некоторые белки плазмолеммы эритроцитов.
В качестве наиболее ярких примеров структурных и транспортных белков ниже будут рассмотрены некоторые белки плазмолеммы эритроцитов.
Как уже упоминалось, в данной мембране – не менее 100 различных белков. Самые известные из них показаны на рис. 14.
Спектрин – фибриллярный белок массой около 240 000 Да, который с помощью другого белка – анкирина – связан с внутренней поверхностью плазмолеммы. Молекулы спектрина имеют вид палочек диаметром 2 нм и длиной около 100 нм. Всего в мембране – более 200 000 таких молекул. Последние на цитоплазматической поверхности мембраны образуют своего рода сетку, которая придает мембране эластичность и упругость. Если в этой сетке молекулы спектрина стыкуются друг с другом конец в конец, то, как показывают расчеты, размер ее ячеек равен примерно 14 нм. Т.е. одна молекула спектрина простирается примерно на 7 ячеек. Если же при контакте друг с другом молекулы спектрина перекрываются, то ячейки, очевидно, будут крупней.
Гликофорин, в отличие от спектрина, не периферический белок, а интегральный. Он пронизывает всю мембрану и с обеих сторон выступает над ее поверхностью. Всего в плазмолемме эритроцита – примерно 360 000 молекул гликофорина.
Белок состоит из двух субъединиц. Каждая из них включает пептидную цепь из 131 остатка аминокислот, а также 16 олигосахаридных цепей, содержащих вместе около 90 остатков моносахаров. При этом на углеводный компонент приходится 60 % массы гликофорина. С этим фактом и связано название белка (буквально: «несущий углеводы»).
В пептидной цепи гликофорина три участка, а именно:
Ø N-концевой участок (~50 остатков аминокислот) выступает с внешней стороны мембраны; с ним-то и связаны все 16 олигосахаридных цепей. Почти каждый второй моносахарид в этих цепях представлен сиаловой кислотой, имеющей отрицательно заряженную группу. Остатки данной кислоты присутствуют также в других белках плазмолеммы; но на гликофорин приходится 75 % от ее общего количества на поверхности эритроцита.
 |
Благодаря сиаловой кислоте, эритроцит несет (на внешней стороне плазмолеммы) значительный отрицательный заряд, который препятствует слипанию эритроцитов друг с другом. Этим эритроциты отличаются от многих других клеток, например нервных и мышечных, у которых внешняя поверхность заряжена положительно.
По мере старения эритроцитов происходит постепенная потеря ими остатков сиаловой кислоты и, соответственно, снижение отрицательного заряда. Когда последний достигает критически низкого уровня, эритроцит захватывается макрофагами селезенки и разрушается.
Таким образом, N-концевой участок гликофорина играет очень важную роль в определении продолжительности жизни эритроцита.
Ø срединный участок (~30 остатков аминокислот) проходит через гидрофобную область липидного бислоя. Вероятно, он имеет вид вытянутой α-спирали, включающей, в основном, аминокислоты с неполярными (гидрофобными) радикалами.
Ø С-концевой участок расположен на внутренней поверхности мембраны и обогащен заряженными и полярными радикалами. Видимо, с ним связываются т.н. актиноподобные белки а с последними, в свою очередь, – элементы цитоскелета клетки (в частности, тонкие микрофиламенты).
В таком случае гликофорин выполняет и очень важную структурную функцию – служит местом крепления цитоскелета.

 Теперь кратко рассмотрим два транспортных белка.
Теперь кратко рассмотрим два транспортных белка.
Белок, образующий анионный канал (масса – 95 000 Да): как и гликофорин, это интегральный гликопротеин, содержащий не менее двух субъединиц. Собственно канал – это узкая пора между субъединицами, «стенки» которой выстланы гидрофильными радикалами аминокислот. Через данный канал в обе стороны (в клетку и из нее) могут проходить анионы (С1-, НСО3-, ОН-) и, по некоторым сведениям, глюкоза.
Заметим, что катионы же через плазмолемму эритроцита почти не проходят: специальных транспортных систем, которые бы пропускали катионы по градиенту концентрации, здесь нет.
Анионные каналы играют важную роль:
ü благодаря им реализуется т.н. эффект Гиббса-Доннана: отрицательно заряженный гемоглобин как бы выталкивает из эритроцитов анионы, отчего их концентрация в эритроцитах оказывается значительно меньше, чем в плазме крови. В частности, из-за меньшей концентрации ионов ОН-, рН в эритроцитах (7,22) ниже, чем в плазме (7,40).
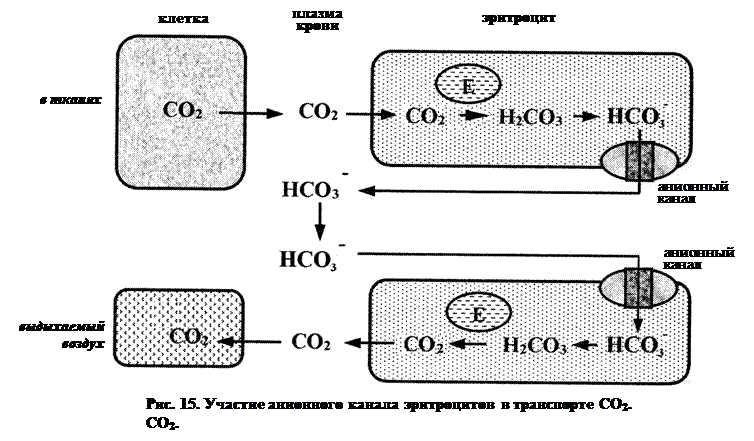
ü анионные каналы связывают плазменный пул бикарбонатных ионов (НСО3-) с карбоангидразой, локализующейся в эритроцитах. В результате, создается единая система переноса СО2 от тканей к легким (рис. 15), где основной транспортной формой являются ионы НСО3-.
Так, в капиллярах тканей СО2 превращается с помощью карбоангидразы в бикарбонат-ионы, которые выходят из эритроцитов в плазму через анионные каналы. В расчете на 1 эритроцит скорость выхода составляет примерно 3,5×108 ионов за 1 с, или ~600 ионов/с для каждого канала. В капиллярах же малого круга кровообращения бикарбонат-ионы, напротив, диффундируют из плазмы в эритроциты, где той же карбоангидразой превращаются в СО2.
Видимо, ввиду важности этих функций, всего в плазмолемме эритроцита оказывается около 600 000 анионных каналов, на которые приходится ~10 % поверхности мембраны и 15% массы всех белков плазмолеммы.
Na+/K+-нacoc (Nа+/К+-зависимая АТФаза) – тоже интегральный гликопротеид. В отличие от предыдущего белка, присутствует в очень незначительных количествах – всего несколько сотен молекул на эритроцит.
О строении и функциях Na+/K+-зависимой АТФазы будет сказано в соответсвующей лекции.
Как и в других клетках, Na+/K+-нacoc за счет энергии АТФ выкачивает из эритроцита ионы Na+ в обмен на ионы К+ и Н+. Но из-за отсутствия К+-каналов (через которые часть ионов К+ выходила бы из эритроцита) на внешней поверхности плазмолеммы не создается положительный заряд, характерный для возбудимых клеток. Поэтому не происходит нейтрализация отрицательного заряда, обусловленного сиаловой кислотой.
|
|
Дата добавления: 2014-11-18; Просмотров: 3132; Нарушение авторских прав?; Мы поможем в написании вашей работы!