КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Опухоли мочевой собирательной системы
|
|
|
|
Вся мочевая собирательная система, начиная от почечной лоханки до мочеиспускательного канала выстлана переходно-клеточным эпителием, или «уротелием». Данный эпителий легко подвергается канцерогенным воздействиям. Этим и объясняется множественность уротелиальных опухолей. Чаще встречается рак мочевого пузыря, несколько реже — опухоли почечной лоханки. Рак мочеточника, как и рак мочеиспускательного канала, встречается редко.
Рак мочевого пузыря. В США рак мочевого пузыря ежегодно диагностируют приблизительно у 40 000 человек; из этого числа 11 000 больных в год умирают. Мужчины болеют в 3 раза чаще, чем женщины. Чаще заболевают люди в возрасте старше 40 лет. Причем замечено, что развитие переходно-клеточного рака преобладает у лиц, подвергающихся воздействию ароматических аминов, особенно 2-нафталамина. Этим, вероятно, можно объяснить высокую частоту появления различных видов уротелиального рака среди курильщиков сигарет и рабочих химических заводов, а также заводов, выпускающих красители и определенные виды резины. Плоскоклеточным раком чаще заболевают лица, у которых выявлена хроническая инвазия Schistosoma haematolium. Возникновение уротелиальных опухолей связывают с длительным введением противоракового алкилирующего средства — циклофосфана, который метаболизируется до активных веществ — акролеина и фосфорамида иприта.
Клинические признаки. Чаще всего больные предъявляют жалобы на макро- и микрогематурию, у них отмечают также дизурию, частое мочеиспускание или недержание мочи. Иногда эти симптомы могут быть единственным проявлением рака мочевого пузыря, и стойкое присутствие их у ранее внешне здорового человека заслуживает внимания. Другие проявления, такие как обструкция мочеточника, боли в области таза или симптомы, связанные с висцеральными или костными метастазами, выявляют при обследовании у небольшого числа больных.
|
|
|
Диагностика и определение стадии процесса. Основными методами диагностики рака мочевого пузыря являются цитологическое исследование мочи, полученной при промывании мочевого пузыря, посредством катетеризации или после мочеиспускания, ВВУ и цистоскопическое исследование с взятием биоптатов из опухоли и выборочно с участков слизистой оболочки, так же как и бимануальное исследование, проводимое под анестезией. К числу выявленных при ВВУ факторов, заставляющих предположить рак мочевого пузыря, относятся односторонняя или двусторонняя обструкция мочеточников с гидронефрозом, дефект наполнения или отсутствие растяжимости мочевого пузыря. Дополнительную информацию относительно стадии процесса можно получить с помощью выполнения КТ брюшной полости или полости малого таза. После эндоскопической резекции опухоли мочевого пузыря оценивают глубину проникновения ее в стенку пузыря. Если дополнительные исследования, направленные на определение стадии заболевания, включая физикальное обследование, рентгеноскопию грудной клетки и биохимический анализ крови, дадут нормальные результаты, то на основании данных биопсии делают заключение относительно того, является ли клиническая стадия заболевания поверхностной или инвазивной. Дополнительную информацию относительно околопузырного распространения опухоли или наличия метастазов в лимфатических узлах можно получить при цистэктомии, которая и укажет истинную патологическую стадию заболевания. У значительного числа больных, стадия заболевания которых отнесена по данным эндоскопии к мышечно-инвазивной, возможны скрытые метастазы в лимфатические узлы или в отдаленные участки тела, если стадию заболевания определяли во время проведения цистэктомии. Такие скрытые микрометастазы указывают на системное поражение, и они ответственны за увеличение числа больных, у которых в конечном итоге возникнут отдаленные метастазы, несмотря на лечение при первичном поражении мочевого пузыря.
|
|
|
Лечение. Рак мочевого пузыря можно подразделить на поверхностный, инвазивный и метастатический. К числу лиц, страдающих поверхностным раком мочевого пузыря, относятся больные с карциномой in situ, с поражением слизистой оболочки (стадия О) или подслизистых слоев (стадия А). Таким больным обычно проводят эндоскопическую резекцию и выборочно берут биоптаты из стенки мочевого пузыря. Через каждые 3—6 мес цистоскопические исследования повторяют. Приблизительно у 50—70% таких больных в течение 3 лет после постановки первоначального диагноза возникает поверхностный рецидив заболевания (поражение слизистой оболочки или подслизистых слоев). Таким больным делают цистоскопическую резекцию и внутрь мочевого пузыря вводят N, N', N''-триэтилентиофосфамид (тиотепа), доксорубицина гидрохлорид, митомицин С или бациллу Кальметта—Герена (БЦЖ).
Еще у 12% больных с начальным поверхностным поражением в конечном итоге разовьется прогрессирующий рак с распространением опухоли в мышцы мочевого пузыря (стадия Б), околопузырную клетчатку (стадия В) или метастазами в лимфатические узлы (стадия Г1), кости или другие внутренние органы (стадия Г2). У некоторых больных первичным может быть инвазивное или метастатическое поражение.
Инвазивное поражение. У больных с инвазивным поражением опухоль проросла в мышцы и/или в околопузырную жировую клетчатку. Традиционными методами лечения считают цистэктомию (радикальная или простая), лучевую терапию или предоперационную лучевую терапию с последующей цистэктомией. При такой схеме лечения приблизительно 45% больных могут прожить еще 5 лет. Причиной летального исхода для большинства таких больных (несмотря на проведение радикальной хирургической операции или лучевую терапию) будут отдаленные метастазы, а не рецидивы местного поражения.
Метастатическое поражение. Из числа больных с отдаленными метастазами в лимфатических узлах, внутренних органах или костях только у 30—70% эффективна системная химиотерапия; но, как правило, улучшение длится не более 6 мес. Большинство больных после появления метастазов живут около 2 лет. Применяемые в настоящее время активные химиотерапевтические средства, такие как цисплатин, метотрексат, доксорубицина гидрохлорид, циклофосфан и винбластин, а также различные сочетания этих средств, обеспечили обнадеживающие долговременные ремиссии. Одна из современных стратегий лечения больных с инвазивным поражением заключается в проведении химиотерапии, а затем осуществлении местного лечения мочевого пузыря (хирургическое или облучение). Целью такой программы является уничтожение микрометастазов, которые обычно имеются у больных с инвазивным поражением.
|
|
|
Переходно-клеточный рак почечной лоханки. Опухоли почечной лоханки составляют приблизительно 10% всех случаев первичного рака почки. Почти 90% из них представляют собой переходно-клеточный рак. Помимо этиологических факторов, вызывающих рак мочевого пузыря, развитию опухолей почечной лоханки способствует нефропатия, вызванная злоупотреблением анальгетиками. Эту форму рака обычно наблюдают у женщин среднего возраста, в анамнезе которых имеются психические заболевания или хронические головные боли и которые в течение нескольких лет приняли большое количество анальгетиков (более 3 кг). Точное количество и вид анальгетиков, вызывающих переходно-клеточный рак, неизвестны, хотя в эксперименте рак этого вида могут индуцировать аспирин и/или фенацетин. Эндемическая балканская нефропатия (см. также гл. 226) связана с высокой частотой случаев развития опухолей почечной лоханки и мочеточников, особенно у женщин.
У большинства больных с переходно-клеточным раком почечной лоханки отмечают безболезненную макрогематурию. Обструкция мочеточника и вызванные тромбами боли встречаются редко. Диагностика основана на результатах ВВУ, с помощью которой выявляют закупоренную, плохо функционирующую или невизуализирующуюся почку или дефект наполнения в визуализируемой почке, и положительных результатах цитологического исследования мочи. Установить природу и локализацию опухоли почечной лоханки или мочеточника позволяют цистоскопия и ретроградная пиелография со взятием биоптата. Консервативное лечение с локальным иссечением пораженного участка и сохранением паренхимы почки на ранних стадиях развития небольших злокачественных опухолей дает благоприятные результаты. После проведенной операции больные живут до 5 лет. При обширных поражениях и на поздних стадиях заболевания лечение заключается в радикальной нефроуретерэктомии и удалении участка мочевого пузыря вместе с мочеточником, идущим от пораженной почки и его устьем. Указанный оперативный подход необходим, чтобы избежать рецидива рака в культе мочеточника и его устье. Кроме того, в дальнейшем обязательно проведение цистоскопического контроля и цитологических исследований мочи чтобы не пропустить возможное впоследствии развитие рака мочевого пузыря и/или опухоли мочеточника или почечной лоханки, расположенных на противоположной стороне. Из числа подобных больных в течение 5 лет выживет 10— 50%. Химиотерапию, разработанную для лечения больных раком мочевого пузыря, применяли и для лечения больных с метастатическим переходно-клеточным раком почечной лоханки, но результаты такого лечения оказались менее успешными.
|
|
|
РАЗДЕЛ 3. ОПУХОЛЕВЫЕ БОЛЕЗНИ
ГЛАВА 293. Т-ЛИМФОТРОПНЫЕ ВИРУСЫ ЧЕЛОВЕКА
Роберт К. Галло, Антоны С. Фаучи (Robert С. Gallo, Anthony S. Fauci)
Биология ретровирусов. Впервые ретровирусы были выделены от цыплят в начале этого века. Позднее, в 50-х годах, от мышей, больных лейкозом, был выделен ретровирус млекопитающего. В настоящее время уже хорошо известно, что эти вирусы связаны с возникновением у многих видов животных как злокачественных, так и незлокачественных заболеваний. Ретровирусы в зависимости от вызываемых ими форм заболеваний могут быть разделены на злокачественные, незлокачественные, одновременно злокачественные и незлокачественные и непатогенные. Непатогенные вирусы часто наследуются как нормальные генетические менделевские элементы. Этот путь их наследования уникален, эти вирусы получили название эндогенных. Примером вируса, обладающего одновременно как злокачественными, так и незлокачественными свойствами, служит вирус, который может вызвать лейкоз у кошек. Он вызывает Т-лейкоз, однако чаще бывает ответствен за нарушения, напоминающие синдром приобретенного иммунодефицита человека (см. гл. 257). Ретровирусы, вызывающие незлокачественные заболевания, например энцефалит и другие неврологические болезни, артриты, болезни легких, относятся к медленно действующим и обозначаются как лентивирусы. Их выделяют от копытных животных, человека (Т-лимфотропный вирус, человека III типа, или HTLV-III) и нечеловекообразных обезьян (обезьяний Т-лимфотропный вирус III типа, или STLV-III).
Ретровирусы имеют внешнюю оболочку, отпочкованную от клеточной мембраны, и содержат электронно-плотную сердцевину, окружающую вирусный геном. Sine qua поп ретровируса представляет собой ДНК-полимеразу, которую называют обратной транскриптазой и которая вместе с РНК входит в состав сердцевины. Обратная транскриптаза катализирует перевод генетической информации из РНК-в ДНК-форму (провирус). Провирус затем обычно мигрирует из цитоплазмы к ядру и после перехода в кольцевую двуспиральную форму интегрируется с ДНК клетки-хозяина, в которой и остается на протяжении всей жизни клетки (рис. 293-1). Поскольку провирус в процессе S-фазы клеточного цикла удваивается вместе с собственно клеточной ДНК, то и дочерние клетки наследуют вирусный геном. Таким образом, организм инфицируется как бы в течение всей жизни. При экспрессии вируса его РНК и белки можно обнаружить в цитоплазме клетки, а также в связи с внутренней поверхностью клеточной мембраны, в которой отпочковывание и высвобождение вируса завершают его жизненный цикл. Иногда происходят делеции провируса, в результате чего формируемый вирус приобретает некоторые свойства, отличающие его от оригинального варианта.
Молекулярные механизмы повреждения клетки вирусом обусловлены структурными особенностями его генома. Наиболее показательны в этом плане вирусы хронических лейкозов, содержащие всего три гена, ответственных за вирусную репликацию: gag, pol и env (рис. 293-2; см. гл. 59). Первый (gag) кодирует синтез внутренних структурных белков, pol — обратной транскриптазы, а env — гликопротеинов оболочки. Свойства оболочки вируса имеют решающее значение в выборе типов клеток, которые он может инфицировать, исходя из чего можно понять требования, предъявляемые к новым антивирусным вакцинам, которые должны способствовать выработке антител, действие которых направлено на оболочку вируса. Вирусная генная цепочка на обоих концах ограничена нуклеотидной последовательностью, которую называют длинным концевым повтором (ДКП) и которая содержит регуляторные элементы, влияющие на экспрессию вирусных генов, а иногда и на вблизи расположенные гены клетки-хозяина. Длинный концевой повтор содержит сигналы, определяющие интеграцию провируса в ДНК клетки-хозяина и образует участки окончания считывания интегрированной провирусной последовательности нуклеотидов. Примерами вирусов, вызывающих хронические лейкозы, могут служить FeLV, вирус лейкоза мышей, вирус лейкоза птиц. До индукции лейкоза эти вирусы интенсивно реплицируются в клетках хозяина. Доказано, что эти вирусы вызывают лейкоз путем интеграции в специфический участок хромосомы, таким образом, ассоциированные ДКП стимулируют постоянную экспрессию клеточных генов, участвующих в процессах роста клетки. Наиболее демонстративным примером этого механизма действия служит развитие у цыплят лейкоза, обусловленного вирусом лейкоза птиц. Его длинный концевой повтор стимулирует экспрессию клеточного онкогена, что, по-видимому, служит первой ступенью в индукции лейкоза. Интеграция вируса носит случайный характер, однако высокая скорость репликации благоприятствует интеграции вируса именно в те участки хромосомы, которые расположены достаточно близко к клеточному онкогену, и, таким образом, ДКП получает возможность его активации. Все это позволяет объяснить совершенно очевидную необходимость длительного размножения вируса до начала развития злокачественного процесса.
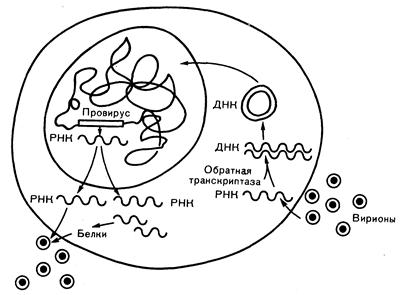
Рис. 293-1. Жизненный цикл ретровируса.
Интактные вирионы проникают в клетку после адсорбции на специфических клеточных рецепторах. Одноцепочечная вирусная РНК лишается оболочки, а обратная транскриптаза синтезирует двуспиральную вирусную ДНК, проникающую в ядро и интегрирующуюся в геной клетки-хозяина. При определенных условиях ДНК провируса не экспрессируется. В других случаях она транскрибируется с образованием РНК, кодирующей вирусные белки, и геномной РНК. Затем происходит сборка вириона из вирусных белков и геномной РНК с последующим его отпочковыванием от клеточной мембраны.
Ретровирус, в результате генетических рекомбинаций приобретающий в своем геноме ген клетки-хозяина, который быстро трансформирует клетки и индуцирует острые злокачественные процессы, часто называют вирусом острого лейкоза или саркомы, а ген обозначают как вирусный onc-ген (см. рис. 293-2; см. гл. 58 и 59).
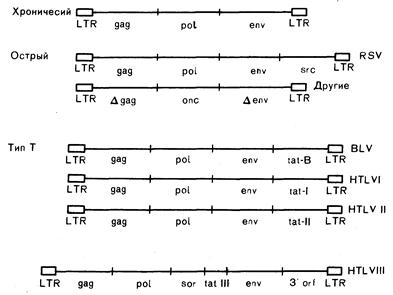
Рис. 293-2. Генная структура и предлагаемая классификация ретровирусов. Синтез внутренних структурных белков кодирует gag, синтез обратной транскриптазы — ро1, гликопротеинов оболочки — env; LTR — длинный концевой повтор; Dgag, Denv — неполные гены; sre — один из онкогенов; BLV — вирус коровьего лейкоза; tat — транскрипционный трансактиватор; sor — короткая открытая рамка считывания; 3'orf—3' — открытая рамка считывания. Функции последних двух генов у HTLV-III не известны.
Вирусы, содержащие onc-ген, немногочисленны и у человека не обнаружены. Они представляют скорее интерес для изучения механизмов неопластической трансформации, чем как этиологический фактор естественного развития опухолей. Каждая клетка, инфицированная этими вирусами, может трансформироваться (развивается поликлональная опухоль), так как продукт вирусного onc-гена оказывает непосредственное трансформирующее действие. Таким образом, для развития опухоли не требуется интеграции вируса в какой-либо определенный участок хромосомы клетки-хозяина.
Общие свойства ретровирусов человека. Третья категория ретровирусов (трансактивационные ретровирусы) представлена хорошо известными вирусами человека: Т-лимфотропными вирусами типов I и II (HTLV-I и HTLV-II) и вирусом коровьего лейкоза (BLV). Третий известный ретровирус человека HTLV-III, или лимфаденопатический вирус, выделен в особую категорию. Геномы всех этих вирусов обладают следующими свойствами: 1) помимо генов вирусной репродукции, они содержат один добавочный ген или более; 2) добавочный ген (гены) негомологичен гену (генам) клеток млекопитающего, т. е. не представляет собой onc-ген; 3) по крайней мере один из добавочных генов кодирует синтез протеина, который участвует в активации экспрессии других вирусных генов и, возможно, некоторых генов клетки (преимущественно за счет связывания с регуляторными усиливающими элементами в клетке, которые сходны с вирусными ДКП). Биологический эффект этих вирусов опосредован именно этим геном, который обозначается как транскрипционный трансактиватор (tat). С учетом того, что tat кодирует синтез протеина ядра, который может активировать другие гены, становится ясно, что для индукции заболевания этому вирусу совсем не требуется интегрироваться в какую-то особую область хромосомы. Следовательно, для развития опухолевого процесса нет необходимости в продолжительной репликации вируса в организме. Сходный феномен отмечается и при развитии лимфомы у коров, вызываемой вирусом BLV. Вирус HTLV-III содержит не только три гена вирусной репликации и ген tat, но еще по крайней мере два других гена, функции которых остаются невыясненными.
Вирусы HTLV-I и HTLV-II сходны по структуре, зрелая форма HTLV-III отличается от них и обладает цилиндрической сердцевиной с высокой плотностью. Первым ретровирусом, идентифицированным у человека, был HTLV-I, который в 1978 г. был выделен от мужчины с агрессивным Т-клеточным злокачественным процессом. Метод идентификации вируса основывался на определении обратной транскриптазы как следа, оставляемого ретровирусом. Этот метод оказался более чувствительным, нежели электронная микроскопия. Помимо этого, с открытием фактора роста Т-клеток, называемого в настоящее время интерлейкином-2, появилась возможность реплицировать вирус in vitro в культуре Т-лимфоцитов-мишеней. Тот же метод использовался и для изоляции вируса СПИДа.
Основное свойство всех известных в настоящее время ретровирусов заключается в их тропизме к Т4 -лимфоцитам (Т-хелперы). Несмотря на то что другие клетки тоже могут инфицироваться этими вирусами, in vitro Т-хелперы в первую очередь повреждаются всеми тремя типами ретровирусов человека; кроме того, при всех заболеваниях, вызываемых ими, практически всегда повреждаются именно эти клетки. Поскольку Т4 -клетки участвуют в регуляции многих иммунных процессов, а также некоторых функций нелимфоидных клеток (см. гл. 62), нетрудно понять, почему эти вирусы вызывают столь серьезные нарушения. Человеческие ретровирусы обладают также способностью имитировать in vitro процессы, протекающие in vivo. Инфицирование Т4+-клеток вирусами HTLV-I или HTLV-II in vitro сопровождается трансформацией некоторых клеток. Свойства трансформированных клеток очень близки к свойствам первично HTLV-1-положительных клеток при Т-клеточном лейкозе взрослых (ТЛВ). Остальные Т4-клетки и другие типы Т-клеток, инфицированные HTLV-I, могут и не трансформироваться, однако у них изменяются некоторые функции. Инфицирование Т4-клеток in vitro вирусом HTLV-III с экспрессией вирусных генов может привести к преждевременной их гибели, что напоминает процессы, протекающие у больных при СПИДе.
Болезни, ассоциированные с HTLV-I. В большинстве случаев лейкозов и лимфом, индуцированных HTLV-I, повреждаются T4+-клeтки, ядро которых приобретает выраженную дольчатость или которые приобретают вид гигантских многоядерных клеток. Однако в ряде случаев не удается выявить каких-либо явных морфологических изменений. Большое значение в патогенезе этих лейкозов имеют существенная экспрессия и увеличение количества рецепторов к интерлейкину-2. Рецепторы к этому фактору роста в течение непродолжительного времени определяются и в здоровых Т-клетках, но только после их иммунной активации. Инфицирование культивируемых интактных Т-клеток сопровождается изменением или полной утратой их иммунных функций. Подобные изменения происходят параллельно развитию оппортунистических инфекций, которые часто сопровождают эти вирусные лейкозы. Лейкозы/лимфомы, вызываемые HTLV-I, обычно протекают в форме лимфоидной неоплазии, известной как Т-клеточный лейкоз/лимфома взрослых (ТЛВ), которой свойственны агрессивное течение, частое развитие гиперкальциемии (механизм не известен), инфекционные осложнения и в половине случаев — формирование лейкозных инфильтратов в коже (см. гл. 294).
HTLV-I может вызывать T4+-клeтoчныe лейкозы/лимфомы, для которых типично более хроническое течение (15—20 % всех случаев), кроме того, они отличаются от ТЛВ и по другим проявлениям. Патогистологически или клинически эти формы заболевания могут быть неотличимы от Т-клеточных хронических лимфоидных лейкозов, диффузных гистиоцитарных или крупно- и смешанноклеточных лимфом, грибовидного микоза или лейкоза Сезари. Лишь у небольшого процента лиц в США эти болезни протекают как HTLV-1-положительный процесс, тогда как практически во всех случаях ТЛВ обнаруживают этот вирус. В некоторых регионах, эндемичных по HTLV-I, определенные В-клеточные лимфоидные и раковые опухоли ассоциированы с HTLV-I чаще, чем это можно было бы ожидать, исходя из распространенности вируса в популяции. В отличие от HTLV-1-положительных Т-клеточных лейкозов, когда вирусные гены интегрированы в ДНК лейкозных клеток,
HTLV-I не обнаруживается в ДНК поврежденных клеток при лейкозах В-клеточного происхождения. Напротив, в этих случаях вирус выявляется в интактных Т-клетках. Малигнизированные В-клетки больных продуцируют единственный тип антител, направленных против протеина HTLV-I. Таким образом, опухоли В-клеточного происхождения образуются (по крайней мере частично) за счет опосредованного влияния HTLV-I, т. е. хроническая антигенная стимуляция, сочетающаяся со снижением иммунных свойств Т-клеток, приводит к увеличению вероятности неопластической трансформации в увеличивающейся популяции В-клеток.
Происхождение и эпидемиология HTLV-I. Несмотря на то что HTLV-I первоначально был обнаружен у двух лиц негроидной популяции в США со спорадической Т-клеточной опухолью, а первые кластеры этого процесcа были выявлены у жителей Японии и несколько позднее у представителей негроидной популяции, уроженцев Карибских островов, этот вирус, по-видимому, имеет африканское происхождение. В пользу этого предположения свидетельствуют: 1) широкое распространение вируса среди населения по всему Африканскому континенту; 2) преимущественная заболеваемость ТЛВ на Американском континенте и в Европе лиц африканского происхождения; 3) обнаружение у африканских примитивных обезьян близкородственного вируса (STLV-I). HTLV-I также широко распространен среди населения двух небольших островов юго-западной Японии (Кюсю и Сикоку), куда он, возможно, был привнесен африканцами в XVI в. Он имеет относительно ограниченное распространение в США и Европе, где инфицировано менее 1 % представителей европеоидной популяции, а у лиц, населяющих Азию, вирус практически не встречается. Таким образом, совсем несложно было установить эпидемиологическую связь между вирусом и вызываемыми им заболеваниями. Передается вирус при половых контактах, переливаниях крови или ее препаратов, а также через плаценту. Выдвигалось предположение о возможном переносе вируса кровососущими насекомыми, однако каких-либо убедительных данных в его пользу получено не было. С увеличением миграции населения, ростом наркомании (использование загрязненных кровью инъекционных игл), изменениями в сексуальном поведении и широким использованием переливаний крови и ее препаратов инфицированность HTLV-I может возрастать.
При инфицировании HTLV-I или HTLV-II небольшая часть Т-клеток становится «бессмертной», теряя потребность в экзогенном интерлейкине-2 для поддержания роста. Этот процесс, по-видимому, опосредован продуктом гена tat, который, как полагают, связывается с регуляторными элементами Т-клеток, активирующих экспрессию гена (генов), участвующего в процессе Т-клеточной пролиферации. Один из этих генов представляет собой рецептор для интерлейкина-2, который, как уже отмечалось, существенно экспрессирован в трансформированных клетках. Причина столь частой трансформации именно Т4+-клеток остается неизвестной, ведь инфицируются не только они, но и другие типы клеток. Поддержание неопластического процесса, вероятно, требует каких-либо дополнительных генетических изменений в клетках, так как гены HTLV-I после развития ТЛВ обычно не экспрессируются.
HTLV-II был первоначально изолирован из культуры клеток, полученных от мужчины, больного Т-вариантом волосатоклеточного лейкоза. Впоследствии вирус был обнаружен еще у двух представителей европеоидной популяции с хроническими формами Т-клеточных опухолей. Получены важные сведения относительно природы HTLV-II генома (он на 50 % гомологичен геному HTLV-I), его эффектов in vitro. Эти два типа вирусов лишь незначительно различаются между собой.
Этиология СПИДа. Этиологическим агентом служит ретровирус, называемый HTLV-III. Этот вирус называют также лимфаденопатическим (LAV) ретровирусом, ассоциированным со СПИДом (ARV). Патогенез синдрома заключается в инфицировании вирусом T4+-индyктopныx/xeлпepныx лимфоцитов, что приводит к преждевременной гибели этих клеток. Наступающий иммунный дефект способствует развитию оппортунистических инфекций и некоторых форм злокачественных опухолей. Детально СПИД обсуждается в гл. 257.
Другие болезни, обусловленные HTLV-III. Помимо характерной картины. СПИДа с оппортунистическими инфекциями, повышенной частотой развития саркомы Калоши и симптомокомплексом, соотносимым со СПИДом (см. гл. 257), инфекция HTLV-III/LAV может быть ассоциирована и с другими заболеваниями. Вирус может инфицировать головной мозг, что приводит к тяжелым нейропсихическим процессам. Известны также случаи лимфоидных интерстициальных пневмонитов, связанных с инфекцией HTLV-III/LAV. При СПИДе повышена заболеваемость определенными формами В-клеточных лимфом. Более того, при инфекции, вызванной HTLV-III/LAV, возможно увеличение частоты заболевания болезнью Ходжкина и некоторыми видами карцином с преимущественной локализацией в области головы и шеи, а также клоакогенной сквамозно-клеточной карциномой. Причина учащения этих злокачественных опухолей еще неясна. HTLV-III/LAV не может служить непосредственной причиной их развития, так как в ДНК большей части опухолевых клеток не удается выявить вирусной нуклеотидной последовательности. В механизме развития В-клеточных лимфом HTLV-III, вероятно, также играет роль опосредующего фактора. При инфекции, обусловленной HTLV-III/LAV, возможно также развитие аутоиммунной тромбоцитопении и наследственных аномалий.
Цитопатическое воздействие HTLV-III/LAV на Т4'-клетки. Инфицирование Т4 -клеток HTLV-III/LAV приводит к их преждевременной гибели. Известно, что гибель инфицированной клетки наступает в результате действия одного или нескольких генов HTLV-III/LAV после внедрения в клетку ДНК-провируса. In vitro индуцировать продуктивное вирусное инфицирование Т4 -клеток невозможно до тех пор, пока они не будут подвергнуты иммунной активации. При активации в инфицированных клетках происходит тот же процесс генной экспрессии, что и в неинфицированных клетках, однако отличие заключается в экспрессии первыми еще и вирусных генов. При этом больший, чем в норме, процент клеток подвергается терминальной дифференцировке, скорость которой выше, чем у интактных клеток. За развитие этих процессов, по-видимому, ответствен ген tat-III (см. рис. 293-2). Экспрессия tat может поочередно значительно усиливать транскрипцию других вирусных генов или же клеточных генов, ответственных за усиление терминальной дифференцировки.
Гетерогенность HTLV-III/LAV. При молекулярном анализе разнообразных изолятов HTLV-III/LAV были выявлены отличия в последовательности нуклеотидов и в определенных участках генома, особенно в гене, кодирующем синтез белков оболочки. Вирусный геном изменяется в процессе последовательного инфицирования клеток, но эти изменения никогда не удается выявить в клетках, культивируемых в течение продолжительного периода. В связи с этим можно предположить, что они происходят при образовании ДНК-транскрипта вирусной РНК и/или рекомбинационном процессе, когда ДНК провируса интегрируется в ДНК клетки-хозяина. Изменения в участках вирусного генома могут в итоге привести к потере вирусной частицей своих инфекционных свойств.
Предупреждение инфекций, обусловленных HTLV-III/LAV, и лечение больных. В предупреждении инфекций, вызванных HTLV-III/LAV, и лечении больных выделяют три проблемы. Во-первых, Т-клетки представляют собой основной тип клеток, ответственных за антивирусные действия, одновременно они же в первую очередь повреждаются вирусом. В целом во всех случаях, когда инфицирование происходит при непосредственном контакте «клетка—клетка», мало что можно сделать для усиления защиты от вируса. Во-вторых, серьезную проблему представляет гетерогенность оболочки вируса в разных изолятах HTLV-III/LAV, однако результаты недавно проведенного сравнения нуклеотидной последовательности, ответственной за синтез белков оболочки, в нескольких изолятах вируса свидетельствуют о том, что они обладают общими сохранными зонами этого гена, некоторые из которых должны быть иммуногенными. Таким образом, теоретически задача создания вакцины для выработки защитных антител представляется вполне разрешимой. В-третьих, поскольку инфицирование означает интеграцию вирусных генов в ДНК клеток, то эти гены передаются потомству поврежденной клетки и, следовательно, инфицирование продолжается постоянно. В США более 1 млн лиц инфицированы HTLV-III/LAV. Важно, чтобы они избегали инфицирования другими агентами, которые могли бы активировать уже инфицированные Т-клетки, что способствует распространению вируса и наступлению смерти. Разработан целый ряд антивирусных препаратов, ингибирующих обратную транскриптазу или воздействующих на оболочку вируса. Другой подход к лечению больных СПИДом основан на данных структурно-функциональных исследований вирусного генома и заключается в создании препаратов, подавляющих функцию или экспрессию гена tat-III. Лечение, по-видимому, следует продолжать в течение всей жизни больного. Для уменьшения токсического действия препаратов и снижения вероятности развития резистентности вируса необходимо применять их комбинации с учетом различий в механизме действия. Еще одним направлением в лечении может быть уничтожение инфицированных клеток. Гипотетически, если можно было бы добиться их полного удаления, то соответственно достигалось бы и полное излечение. На практике, однако, осуществить это невозможно, поскольку большая часть инфицированных клеток не экспрессирует вирусные протеины и, следовательно, ничем не отличается от здоровых клеток.
СПИД представляет собой относительно новое инфекционное заболевание человека и характеризуется развитием тяжких осложнений, часто приводящих к смерти больного. Подобно инфицированию HTLV-I (и, вероятно, HTLV-II), инфицирование вирусом СПИДа лиц, проживающих в Африке, произошло от зеленых мартышек или родственных видов обезьян при прямом контакте либо опосредованно через промежуточных переносчиков с последующим распространением на лиц, проживающих в других регионах. HTLV-III/LAV сходен и с HTLV-I по механизму передачи, тропизму к Т4 -клеткам, поведению in vitro и гену tat. В отличие от HTLV-I и HTLV-II вирус СПИДа содержит по меньшей мере еще два дополнительных гена, оказывает выраженное цитопатическое действие, отличается большим структурным сходством с лентиретровирусами и в целом более высокой инфекционностью.
ГЛАВА 294. БОЛЕЗНЬ ХОДЖКИНА И ЛИМФОЦИТАРНЫЕ ЛИМФОМЫ
Винсент Г. ДеВита, Джон Е. Ултман
Определение. Лимфомы следует рассматривать как опухоли иммунной системы. К ним относятся лимфоцитарные опухоли и болезнь Ходжкина, а иногда в группу лимфом включают и опухоли гистиоцитарного происхождения. Ранее лимфомы подразделяли на болезнь Ходжкина и неходжкинские лимфомы, но в настоящее время более совершенные методы диагностики позволяют определеннее высказаться о характере заболевания. Старые термины употреблять не следует.
Эпидемиология. В 1985 г. было зарегистрировано 34 000 новых случаев заболевания лимфомами, 40 % из которых приходилось на болезнь Ходжкина. Наиболее распространенными среди лимфоцитарных лимфом бывают фолликулярные лимфомы, а также диффузные крупноклеточные лимфомы. На каждую из этих форм приходится примерно 40 % от всех лимфом, за исключением болезни Ходжкина. Средний возраст больных относительно небольшой (32 года при болезни Ходжкина и 42 года при всех остальных формах лимфом взрослых), и по показателю непрожитых человеко-лет лимфомы занимают 4-е место среди всех опухолевых заболеваний. Несмотря на то что заболеваемость лимфомами год от года повышается, в результате совершенствования методов лечения уровень смертности при них постоянно снижается. Например, с 1970 г. продолжительность жизни при болезни Ходжкина существенно увеличилась, и с 1973 г. по настоящее время общенациональный показатель смертности в США уменьшился на 58 %. При диффузных крупноклеточных лимфомах общенациональный уровень 5-летнего срока жизни повысился с менее чем 5 % до 40 % и более.
Больные с лимфомами распределяются по регионам мира неравномерно. В США отмечают два возрастных пика заболеваемости лимфомой Ходжкина, один из которых приходится на возраст 15—35 лет, а второй — на возраст после 50 лет. У молодых заболевание особенно часто протекает по типу нодулярного склероза. В Японии ранний возрастной пик не определен. Болезнь Ходжкина у детей в возрасте до 10 лет регистрируется преимущественно в слаборазвитых странах, при этом ее гистологические формы и распространенность соответствуют таковым при наиболее запущенных случаях. Все это, а также немногочисленные сообщения об очагах распространения болезни позволяют думать о роли окружающей среды и/или генетических факторов в ее развитии. После сообщения о том, что в США выявлено несколько очагов болезни Ходжкина, были проведены популяционные исследования с использованием раковых регистров штатов Коннектикут и Калифорния. Результаты этих исследований свидетельствуют о том, что в этих случаях дело имели, вероятно, со случайными заболеваниями. Медицинский персонал, контактирующий с больными, заболевает не чаще, чем любой человек в популяции. Однако результатами тщательно проведенного эпидемиологического анализа доказано, что болезнь Ходжкина, возможно, представляет собой необычное проявление какой-то распространенной инфекции. Установлено, что факторы, повышающие риск раннего контакта с инфекциями (например, большие семьи, скученность проживания нескольких семей), одновременно снижают риск развития болезни Ходжкина. Эти данные свидетельствуют и о том, что, по-видимому, у лиц молодого и пожилого возраста в развитии болезни играют роль разные факторы риска, чем можно объяснить некоторые курьезные результаты эпидемиологических наблюдений, например тот факт, что пик заболеваемости в раннем возрасте в Японии не зарегистрирован. Уникальные эпидемиологические характеристики свойственны некоторым формам лимфоцитарных лимфом. Так, лимфома Беркитта распространена преимущественно среди детей в странах Центральной Африки, в то время как в США зарегистрированы единичные случаи заболевания, причем с другими клиническими проявлениями. Абдоминальные лимфомы, продуцирующие иммуноглобулины с тяжелыми цепями, регистрируются преимущественно в странах Средиземноморья, в других же регионах мира они крайне редки.
Источники лимфом. Лимфомы развиваются в лимфатических узлах или в лимфоидных тканях паренхиматозных органов (кишечник, легкие), а также в коже. У 90 % лиц с болезнью Ходжкина первично повреждаются лимфатические узлы, а у остальных 10 % источником заболевания служит внеузловой очаг. Первичная локализация процесса в паренхиматозных органах при лимфоцитарных лимфомах встречается чаще: 60 % лимфом происходят из лимфатических узлов, а 40 % — из опухолевых очагов другой локализации.
Таблица 294-1. Клеточное происхождение злокачественных лимфом
| Опухоли | ||
| В-клеточные | Т-клеточные | гистйоцитарно/ретикулярно-клеточные |
| Хронический лимфоидный лейкоз, 98 % | Хронический лимфоидный лейкоз, 2 % | Злокачественный гистиоцитоз (гистиоцитарный медуллярный ретикулёз) |
| Малая лимфоцитарная (хорошо дифференцированная) лимфома | Грибовидный микоз/синдром Сезари | Моноцитарный лейкоз |
| Промежуточная и/или малая расщепленная лимфоцитарная лимфома | Диффузные агрессивные лимфомы взрослых, 25 % Смешанные Большие, иммунобластные | Крупноклеточные лимфомы, <5 % Болезнь Ходжкина |
| Фолликулярные лимфомы | Т-клеточный лейкоз/лимфома взрослых | |
| Диффузные агрессивные лимфомы взрослых, 65% | Ангиоцентрические лимфомы (лимфоматозный гранулематоз) (полиморфный ретикулёз) | |
| Смешанные | ||
| Большие | ||
| Большие иммунобластные | ||
| Малые нерасщепленные | ||
| Лимфома Беркитта (малая нерасщепленная) | ||
| Острый лимфоидный лейкоз, 70 % | Острый лимфоидный лейкоз, 25 % | |
| Лимфобластные лимфомы, 10 % | Лимфобластные лимфомы, 85 % |
Фенотип. С появлением более специфичных сывороток появилась возможность классифицировать лимфомы в зависимости от их клеточного происхождения (табл. 294-1). В 75 и 30—40 % случаев лимфомы лимфоцитарного происхождения обусловлены соответственно моноклональными В- и Т-клеточными популяциями. С помощью зондов ДНК к гену иммуноглобулина и b-цепи Т-клеточных рецепторов можно определить принадлежность лимфоидной опухоли к Т- и В-клеточной линиям. Однако при использовании этих методов в ряде случаев было установлено, что до 10 % опухолей имеют биклональную природу, т. е. обладают признаками как Т-, так и В-клеток. Несмотря на морфологическое сходство клеток злокачественных лимфом с тканевыми гистиоцитами, лишь немногие из них, по-видимому, представляют собой истинные производные последних. Помимо этого, есть возможность отнести В-клеточную опухоль к тому или другому функциональному подтипу В-клеточной популяции. Например, фолликулярные лимфомы представляют собой производные пролиферативного очага В-системы — лимфоидного фолликула, тогда как диффузные малые лимфоцитарные лимфомы развиваются из секреторных компонентов мякотных тяжей. Лимфомы В-клеточной природы распознаются по моноклональному иммуноглобулину, их поверхности, а при его отсутствии — по реаранжировке генов иммуноглобулинов при помощи специфических зондов ДНК. В США реже, чем в других странах, встречаются больные с лимфомой Т-клеточного происхождения. Примерно в 15—35 % случаев диффузные большие (крупноклеточные) лимфомы имеют Т-клеточную природу, поэтому их называют периферическими Т-клеточными в отличие от незрелых Т-клеточных тимического происхождения (например, лимфобластная лимфома у детей и подростков). Т-клетки идентифицируют по характерной для них способности образовывать розетки с эритроцитами барана, а также по взаимодействию с моноклональными антителами к дифференцировочным антигенам. Иногда исследование фенотипической экспрессии Т-клеток дополняется изучением их функциональных свойств. При грибовидном микозе/ синдроме Сезари, представляющем собой периферическую Т-клеточную лимфому, клетки несут маркеры хелперов. У этих же клеток определяют и соответствующие функциональные свойства. Для характеристики Т-клеток получено большое количество моноклональных антител к их дифференцировочным антигенам. К специфическим В-клеточным антигенам антител получено меньше. Наибольшей специфичностью обладают моноклональные антитела анти-В1 (пан-В-клеточные антитела). Широко используются и другие моноклональные антитела (I5), которые, как первоначально предполагали, реагируют исключительно с общим антигеном, ассоциированным с острым лимфобластным лейкозом (CALLA). В дальнейшем было показано, что этот антиген экспрессируется при многих В-клеточных опухолях, включая большинство фолликулярных лимфом и лимфому Беркитта. Источником болезни Ходжкина могут быть дендритные ретикулярные клетки, осуществляющие презентацию антигена и локализующиеся в паракортикальных зонах лимфатических узлов. В культуре клетки Штернберга—Рид и их мононуклеарные варианты несут на мембране Ia-антигены, Fc и С3-рецепторы. Они не синтезируют иммуноглобулины, неспособны к фагоцитозу, в них не выявляется диффузная активность неспецифической эстеразы и кислой фосфатазы. Все это свидетельствует в пользу происхождения клеток Штернберга—Рид от антигенпрезентирующих клеток. Клетки Штернберга—Рид могут образовывать розетки с Т-клетками и даже в парафиновых срезах реагировать с моноклональными антителами анти-Leu Ml, также взаимодействующих с дендритными ретикулярными клетками после обработки их нейраминидазой. Антитела анти-Leu Ml не реагируют с морфологически сходными Т-клетками. Моноклональные антитела Kil, направленные против клеточных линий болезни Ходжкина, также взаимодействуют с этими клетками в замороженных срезах лимфатических узлов, однако, как было установлено, они реагируют и с поверхностными антигенами при больших (крупноклеточные) лимфомах В-клеточного происхождения. Ряд моноклональных антител используется для определения общих лейкоцитарных антигенов, т. е. экспрессируемых на всех нормальных лимфоретикулярных клетках. Эти антигены позволяют дифференцировать карциномы и саркомы от злокачественных лимфом.
Этиология. Существуют доказательства того, что вирусы могут вызывать лимфомы у грызунов, птиц, кошек и коров. Впервые вирусная природа лимфомы была доказана американскими и японскими исследователями, которые выделили уникальный ретровирус от больных грибовидным микозом в США и острой Т-клеточной лимфомой в Японии. Последнее заболевание относительно новое, редко встречающееся в США, но весьма часто в Японии и среди лиц негроидной популяции, проживающих в странах Карибского региона. Этот класс вирусов назвали вирусом человеческого Т-клеточного лейкоза/лимфомы (HTLV). По этой причине предполагают, что и другие лимфомы человека могут быть обусловлены этим вирусом. Отмечены случаи развития диффузных иммунобластных лимфом у больных с синдромом приобретенного иммунодефицита (СПИД). Несмотря на то что до сих пор HTLV идентифицируют как Т-лимфотропные вирусы, обнаружен дальнеродственный вирус коровьей лимфомы/лейкоза, вызывающий В-клеточный лейкоз/лимфому. Вирус, подобный HTLV, был изолирован также из клеток В-клеточных лимфом человека. В настоящее время с помощью современных методов культивирования клеток лимфом в длительно поддерживаемых культурах вирусная этиология лимфом человека проверяется. Вирус может рассматриваться как этиологический фактор болезни Ходжкина, при которой нарушения иммунитета, сопровождающие заболевание, имеют Т-клеточную основу и могут быть следствием инфицирования Т-клеточным лимфотропным вирусом. Кроме того, отмечена строгая связь между ДНК-содержащим вирусом Эпстайна—Барр (ВЭБ) и редкой лимфомой, охарактеризованной Беркиттом, обследовавшим больных в Восточной Африке (в их сыворотке обнаруживают антитела к ВЭБ, а в человеческом геноме клеток Беркитта определяют комплементарную ДНК). Связь лимфомы Беркитта с ВЭБ у больных, проживающих в США, представляется менее постоянной. Помимо этого, у большого числа лиц, перенесших инфекционный мононуклеоз (вызываемый ВЭБ), по прошествии продолжительного периода незначительно, но постоянно повышается уровень заболеваемости лимфомами по сравнению с контрольной группой лиц. Другая разновидность герпесподобного ДНК-содержащего вируса вызывает лимфоматоз цыплят (болезнь Марека), для профилактики которой в настоящее время создана вакцина.
Значение наследственных факторов в развитии лимфом подчеркивается повышенной частотой заболеваемости среди лиц с наследственными формами иммунной недостаточности и незначительным увеличением заболеваемости среди членов семей больных с иммунными нарушениями. Как было отмечено, у потомства родителя, страдающего болезнью Ходжкина (особенно у однополого), риск заболевания резко повышается. Несколько чаще лимфомы сопровождают коллагенозы у больных сопоставимого возраста. Увеличение их частоты составляет 10% при длительно текущем синдроме Шегрена, при котором отмечается склонность к развитию диффузных лимфом или иммунобластных сарком.
Лимфоподобные синдромы появляются у лиц, принимающих фенитоин. Несмотря на то что в большинстве случаев болезненные проявления разрешаются после прекращения приема препарата, у значительного числа лиц развиваются истинные лимфомы нескольких разновидностей, включая болезнь Ходжкина. По-видимому, действие фенитоина объясняется генетической предрасположенностью к лимфомам. Частота диффузных крупноклеточных и иммунобластных лимфом (часто с вовлечением в процесс головного мозга) повышена при состояниях хронической иммунодепрессии, особенно при синдроме приобретенного иммунодефицита и после трансплантации почки или сердца.
Цитогенетические нарушения при лимфомах. Опухолевые клетки при болезни Ходжкина проявляют анеуплоидные свойства, однако каких-либо специфичных хромосомных аномалий при этом заболевании не установлено. Напротив, неслучайные хромосомные изменения свойственны практически всем формам лимфом, причем чаще всего в процесс транслокации вовлекается хромосома 14 (8; 14, 11; 14, 14; 18). Впервые транслокация была выявлена при лимфоме Беркитта. В ее результате онкоген c-myc, локализующийся в нормальной позиции на хромосоме 8, оказывается в непосредственной близости к промоторной последовательности гена, кодирующего тяжелые цепи иммуноглобулина, в результате чего усиливается экспрессия c-myc. Другие частые транслокации (8; 12, 8; 22) переводят тот же онкоген под контроль промоторов соответственно -л- и \- цепей. Транслокации 14; 11 и 14; 18 свойственны фолликулярным лимфомам, причем обнаружены два ранее не известных гена, вероятно, представляющих собой лимфомоспецифичные онкогены, локализующиеся в хромосомах 11 и 18 (обозначены как BCL1 и BCL2) в непосредственной близости к точке разрыва. Метод клонирования рекомбинантной ДНК, содержащей фрагменты ДНК из области точек разрыва при этих транслокациях, позволил разработать специфические зонды, которые могут служить средством идентификации лимфоцитов с этими хромосомными нарушениями.
|
|
|
|
|
Дата добавления: 2014-11-20; Просмотров: 450; Нарушение авторских прав?; Мы поможем в написании вашей работы!