Теория лимитов популяционной численности, или теория X. Андреварты — Л. Бирча
Теория (принцип) биогеоценотической (экосистемной) регуляции численности популяции К. Фридерихса
Правило пищевой корреляции В. Уини-Эдвардса Теория (принцип) Д. Лэка
Правило стрессогенного увеличения надпочечников Я. Кристиана
Теория общего адаптационного синдрома Г. Селье
Теория Дж. Кристиана — Д. Дейвиса
Правило сохранения видовой среды обитания
Правило колебаний (цикличности) численности
Правило максимума размеров колебаний плотности популяционного населения, или закон количественной константности популяционного населения
Правило максимальной рождаемости (воспроизводства)
Правило стабильности половозрастной структуры популяции
Принцип территориальности
Принцип скопления (агрегации) особей В. Олли
Правило топографического, или популяционного, кружева ареала
Принцип стабилизации экологической ниши, или принцип биоценотической коэволюции
Принцип, или теорема В. Людвига
Теория (принцип) А. Николсона
* Многие из них так или иначе затронуты в книге: А. М. Гиляров. Популяционная экология. М.: Изд-во МГУ, 1990. 191 с.
Число сформулированных биоэкологами популяционных закономерностей очень высоко — до 20, а быть может, значительно больше*. Это понятно — все живые существа так или иначе входят в популяционные объединения. Если эти объединения рассматривать не только как функциональные, но и пространственные образования, т. е. биогеографически, то, пожалуй, число закономерностей возрастет еще больше. Поэтому есть смысл разделить собственно популяционные и биогеографические обобщения. Кроме того, часть закономерностей, обычно относимых к популяционным, рассмотрены в разделе о функционировании сообществ (3.8) или освещены ранее (разд. 3.4) при разборе эколого-организменных взаимосвязей (например, принцип внезапного усиления патогенности) и взаимодействий в системах организм — среда (разд. 3.5). Вообще разделение подобных групп закономерностей весьма условно: популяция состоит из особей и не существует изолированно от среды; то же утверждение относится к любым другим образованиям с участием живого. Вероятно, наиболее общим правилом является то, что индивиды любого вида живого всегда представлены не изолированными отдельностями, а их определенным образом организованными совокупностями. Это правило получило имя С. С. Четверикова, сформулировавшего его в 1903 г. Его можно назвать правилом объединения в популяции. В связи с тем, что любая популяция обладает строго определенной генетической, фенотипической, половозрастной и другой структурой, она не может состоять из меньшего числа индивидов, чем необходимо для обеспечения стабильной реализации этой структуры и устойчивости популяции к факторам внешней среды. Именно в этом состоит принцип минимального размера популяций. Такой размер не есть константа для любых видов, он строго специфичен для каждой популяции. Выход за пределы минимума чреват для популяции гибелью: она уже не будет в состоянии самовозобновиться. Однако ее возможно восстановить искусственно, поскольку каждый индивид содержит в себе хотя и обедненное, но все же весьма значительное число генетических задатков вида, к которому он принадлежит.
* Одум Ю. Основы экологии. М.: Мир, 1975. С. 255. ** Асимптота — прямая линия, к которой неограниченно близко стремятся точки некоторой кривой по мере того, как эти точки удаляются в бесконечность; асимптотический — неограниченно приближающийся.
Естественно предположить, что если есть минимум, то возможен и максимум. Это предположение помимо логической посылки основывается на соотношении законов максимума биогенной энергии и давления среды, упомянутых в разд. 3.5.1. Ю. Одум формулирует закон как правило популяционного максимума в следующих словах*: «Они (популяции — Я. Р.) эволюционируют так, что регуляция их плотности (населения — Я. Р.) осуществляется на значительно более низкой по сравнению с верхней асимптотой** емкости местообитания, достигаемой лишь в том случае, если полностью используются ресурсы энергии и пространства». Этому утверждению предпосылается фраза: «В 60-е годы получены данные, показывающие, что дело вовсе не в тенденции популяций избегать гибельных отклонений в обе стороны от уровней насыщения». Высказывание несколько странное, поскольку речь идет именно о саморегуляции численности популяции, вернее, плотности ее населения. Общая же абсолютная численность особей популяции ограничена произведением максимума плотности населения на жизнепригодную для популяции площадь обитания с учетом разницы в местообитаниях. Ю. Одум на той же странице приводит данные В. Уини — Эдвардса (1962 и 1965 гг.) о двух механизмах, управляющих системой превентивного самоограничения плотности населения: 1) территориальном поведении особей и их малых групп как форме внутривидовой конкуренции и 2) групповом поведении как сбалансированности противоположных форм активности (конкуренция и кооперация, агрессивность и подчинение при этологической иерархии и т. п.). О межвидовых отношениях в процессе генетической обратной связи в популяциях лишь упоминается со ссылкой на работы Д. Пиментела. Правило популяционного максимума конкретизируют два обобщения. Первое из них известно как теория лимитов популяционной численности, или теория X. Г. Андреварты — Л. К. Бирча (1954): численность естественных популяций ограничена истощением пищевых ресурсов и условий размножения, недоступностью этих ресурсов и слишком коротким периодом ускорения роста популяции. Второе обобщение дополняет первое. Оно носит название теории биоценотической регуляции численности популяции К. Фридерихса (1927): регуляция численности популяции есть результат комплекса воздействий абиотической и биотической среды в местообитании вида. Совершенно очевидно — теория К. Фридерихса объясняет, что лимитирует и формирует потенцию роста численности популяции, заложенную генетически. Однако, как известно, бывают взрывы роста численности популяций — сбои в регуляторных системах самоограничения и внешних ограничений популяционного роста. Несколько уже, чем у Ю. Одума и в упомянутых теориях, подход к взаимосвязи популяции и среды ее обитания у В. Уини-Эдвардса (в ряде случаев пишут Вини-Эдвардс), который сформулировал правило пищевой корреляции: в ходе эволюции сохраняются только те популяции, скорость размножения которых скоррелирована с количеством пищевых ресурсов среды их обитания. Важно подчеркнуть, что скорость размножения в норме всегда ниже максимально возможной, и постоянно остается запас пищевых ресурсов. Очевидно, что размер видового населения в ходе эволюции должен быть ограничен не текущими пищевыми возможностями, а их минимумом, возникающим или могущим возникнуть в течение достаточно большого ряда лет. В противном случае популяция столкнется с резким недостатком пищи и в значительной своей части вымрет как под воздействием прямого недостатка кормовых объектов, так и в результате межвидовой конкуренции, осложняемой биоценотическими регулирующими механизмами в целом (к этому вопросу мы еще вернемся в конце параграфа при обсуждении принципа стабилизации экологических ниш). Для того чтобы избежать подобной коллизии, и была эволюционно выработана преадаптация более низкой плотности населения популяции, чем допускают наличные кормовые возможности. Конечно, при обсуждении проблем саморегуляции ограничивать действие регулирующих факторов среды одними лишь условиями питания нельзя. Следует говорить о емкости среды в целом, как это сделал Ю. Одум. Однако кормовые возможности играют одну из ведущих ролей и прежде всего бросаются в глаза. Их легче рассчитать математически. Несколько шире ставит вопрос Д. Лэк, сформулировавший в 1954 г. принцип, или теорию, носящую его имя, теория, или принцип Лэка: среднее количество яиц, откладываемых птицами, эволюционно ограничено возможностями выращивания выведшихся из них птенцов. Очевидно, этот лимит связан с комплексом условий: интенсивностью обмена веществ, длиной дня, наличием пищи, ее близостью и т. д. В основе, безусловно, лежит энергетика системы — она отработана эволюцией вида в конкретных условиях его обитания*. Энергетическая расточительность в природе слишком большая роскошь, хотя бывают и исключения.
* В более обобщенном виде вопрос о репродуктивных ограничениях сформулирован в виде правила максимальной рождаемости (воспроизводства) — см. ниже этот же раздел. ** Некоторые культуры исторически выработали механизмы самоограничения, например, тибетцы. Был широко распространен инфантицид, особенно девочек. У северных народов эволюционно сниженная плодовитость, но основная масса народов мира не имеет механизмов демографического самоограничения, и регуляция шла главным образом за счет повышенной смертности. При сверхплотном населении городов у женщин не происходит спонтанного рассасывания эмбрионов и других процессов, характерных для животных. Впрочем, и эти процессы не предохраняют многие виды (например, лосей) от сверхразмножения с полным уничтожением кормовой базы. Демографическое регулирование у людей может основываться лишь на специальных мероприятиях. К этому вопросу мы вернемся в последних главах книги.
Хорошо известно, что в ряде случаев происходят массовые размножения организмов, не предотвращаемые ни внутрипопуляционными, ни межвидовыми механизмами. При этом не срабатывает система эволюционной взаимосвязи: плотность населения животных как. фактор отбора для растений — изменение генетической конституции растений, а также обратная связь: генетическая конституция растений — плотность населения животных. К тому же популяционные механизмы саморегуляции в мире животных никак нельзя переносить на популяции растений. У последних нет ни территориального, ни группового поведения, аналогичного тому, что существует у животных. При переходе к человеческому обществу приходится констатировать, что биологическая превентивная саморегуляция численности у него, как правило, отсутствует, а социально тысячелетиями доминировало стремление «плодитесь и размножайтесь»**. Иначе бы в истории людей не возникли локальные, региональные и глобальные экологические кризисы. Очевидно, мы имеем дело с некой эволюционной нормой саморегуляции и закономерными отклонениями от нее. В растительном мире, как правило, наблюдается максимальное заполнение пространства и использование солнечной энергии при сохранении запасов минерального питания. Лимитирующими факторами служат физическая заполненность территории, поток лучевой энергии, предшествующие события, материализованные в процессах почвообразования, собственное воздействие растений на среду (например, аллелопатия) и сложные взаимосвязи с представителями других царств природы — микроорганизмами, грибами и животными. Превентивных механизмов саморегуляции плотности населения не наблюдается. Однако существует мощный рычаг ее восстановления в случае изреживания — почвенный запас семян, спящие почки и т. п. И параллельно этому имеется генетический механизм ограничения размеров индивидов. В противном случае одно растение было бы способно занять всю площадь и не оставалось бы места для процессов дублирования как инструмента обеспечения надежности системы. Емкость местообитания используется растениями максимально полно в рамках совокупности экологических ограничений. Основной их биотический механизм перенесен в область взаимодействия собственных размеров, межвидовых отношений и индивидуального воздействия на среду жизни. Свободный запас вещества и энергии минимален. В основном действуют факторы, непосредственно зависящие от плотности населения популяции. Однако по отношению к минеральному питанию, т. е. к «пище» растений, как было упомянуто выше, они ведут себя подобно животным, имея механизм эволюционной преадаптации, препятствующий истощению почв. Этот механизм ориентирован на скорость почвообразовательных и шире — биогеоцентрических процессов. В одних случаях запас велик (например, черноземы), в других он очень мал (тропические леса). Чем интенсивнее круговорот веществ в экосистеме, тем ниже положительный баланс соотношения ресурсов минерального питания растений и скорости роста их популяций. Растения «берут в основном числом, а не умением».
* По массе надземной части, по энергетике обычно не более 1 %. С точки зрения энергетических «запасов» растительность не выходит за рамки этого же правила — используется не более 10 % поступающей солнечной энергии, чаще всего лишь 0,2 ее процента. Здесь говорится только о веществе.
В царствах дробянок, грибов и животных, как правило, существует иная закономерность: потенциальный запас жизненных ресурсов огромен — обычно используется не более 30 — 60% наземной луговой растительности (крупные млекопитающие) и 10% лесной растительности*. Более интенсивно и полно утилизируется растительноядными животными чистая продукция водных растений. Как хорошо известно, пирамида биомасс в водной среде как бы перевернута — продуценты размножаются столь интенсивно, что консументы могут превышать их по биомассе. Однако это касается лишь мелких обитателей водной среды. Для рыб, птиц и млекопитающих общая их биомасса всегда намного ниже, чем потребляемых ими растительных кормов. Случаи массового размножения организмов следует отнести к двум группам. Одна из них связана с антропогенными трансформациями среды жизни. Например, в сибирской тайге очаги массового размножения вредителей никогда не возникают в коренных лесах. Другая группа, видимо, связана с особым типом отношений между популяционной динамикой и экосистемными механизмами. Эволюционный отбор идет не на уровне внутрисистемных отношений в рамках собственной экосистемы, а на межэкосистемном уровне. Таковы вылеты саранчи, некоторых бабочек, отчасти миграции леммингов, белок, из птиц — кедровок. В первом случае массовое размножение стимулируется аномальными для популяции условиями жизни: повышением потока энергии сверх обычного уровня, необычными условиями питания, снижением пресса конкурентов, хищников и паразитов, особо благоприятными условиями гидрологического режима и т. п. Переразмножение в изначальном очаге перебрасывается на широкие площади и затем резко обрывается с гибелью значительной части популяции от недостатка кормов или с развитием болезней и пресса других врагов. Во втором случае арена действия принципиально схожа, но значительно пространственно шире. Аналогия в том, что вспышка массового размножения возникает в одних экосистемах и перекидывается на другие. Но имеется существенная разница. Вместо узких очагов переразмножения — обширное его поле. Вместо локального воздействия — широко межрегиональное, значительные выселения особей (пешая саранча, летные ее кулиги, переселения, часто безвозвратные, млекопитающих и птиц). Как и в первом случае, затухание происходит как в изначальном ареале массового размножения, так и вне его, но в первом случае в сходных с очагом размножения экосистемах, а во втором часто — в резко отличных. И в этом втором случае действуют более высокие по иерархии экосистемы, «тушащие популяционный пожар».
* Тут следует сослаться на огромное число работ, опубликованных исследователями из морфофизиологической школы С. С. Шварца. Перечислить эти работы нет никакой физической возможности.
Здесь уместно упомянуть одно частное правило, а именно правило стрессогенного увеличения надпочечников у высших позвоночных животных Я. Кристиана. Оно было «канонизировано» в 1963 г. уже после широкого практического использования этой закономерности*. Заключается оно в том, что в условиях перенаселенности у высших позвоночных животных увеличиваются надпочечники. Это результат сдвига нейроэндокринного гомеостаза, ведущего к изменению поведения животных (в частности, к повышению их агрессивности), репродуктивного потенциала (происходит резорбция эмбрионов), снижению устойчивости к заболеваниям и другим последствиям. Комплекс этих изменений обычно ведет к резкому падению жизнеспособности особей, их массовой гибели, снижению плотности населения популяции или даже ее элиминации от «стрессового шока». Как ясно из сказанного, основой обсуждаемого правила служит теория общего адаптационного синдрома Г. Селье. Частное наблюдение стрессогенного увеличения надпочечников вкупе с теорией общего адаптационного синдрома Г. Селье вылилось в теорию Дж. Кристиана — Д. Дейвиса, которые в работах 1950—1968 гг. объяснили регуляцию численности популяций млекопитающих их социальным поведением, отражающимся на эндокринных реакциях (что и ведет к увеличению надпочечников). У нас в стране эту точку зрения активно пропагандировал С. С. Шварц, создавший большую школу исследователей этого направления. Их привлекала прежде всего простота получения эмпирического материала.
* Значительная часть из них упомянута в кн.: Реймерс Н. Ф. Птицы и млекопитающие южной тайги Средней Сибири. М.-Л.: Наука, 1966. 420 с. ** К сожалению, на мои работы никто не обратил внимания — ни за рубежом, ни внутри страны. У нас просто не было социально-научной основы для «ассимиляции» экологических знаний, а за рубежом русскоязычные работы мало доступны. Кроме того, в советских журналах статьи ждали публикации по 3 — 4 года.
В цикле работ 50-х — начала 60-х гг.*, видимо, одновременно с В. Уини-Эдвардсом, мне** пришлось показать, что позвоночные, да и беспозвоночные животные в норме не могут абсолютно разрушительно воздействовать на среду своего обитания (правило сохранения видовой среды обитания), поскольку такое разрушение лишает их собственной основы жизни. Грубо говоря, съесть сразу все означает затем остаться без пищи и погибнуть от голода. Естественно, эволюционно должны были возникнуть ограничивающие механизмы популяционного и внепопуляционного характера (последние объединяются правилом относительной внутренней непротиворечивости, см. разд. 3.8.3). С одной стороны они обеспечивают возможность поиска (или «поиска» для неживотных) корма, с другой — ограничивают такой поиск или снижают саму плотность населения через популяционные механизмы — физико-химические и поведенческие. Для нас сейчас не столько важны эти механизмы, как сама констатация того, что в нормальных условиях действует закон (правило) популяционного максимума и правило пищевой корреляции, имеющее множество модификаций в зависимости от среды жизни (водной, наземной и т. д.) и от царства природы, даже систематического типа и класса животных (членистоногие, млекопитающие и т. д.). Группа закономерностей саморегуляции в системах живого имеет важное практическое и теоретическое значение. Виды (популяции) поддерживают собственную среду жизни, то же происходит в рамках биоценозов и других биотических образований, в том числе на уровне всего живого вещества планеты — см. правило автоматического поддержания глобальной среды обитания (разд. 3.10). Если механизмы самоподдержания на популяционном уровне относительно независимы от деятельности человека, то деформируя биоценозы, человек сильно видоизменяет гомеостаз на уровне ценозов. Разрушение этого механизма в биосфере означало бы глубокую трансформацию жизни или даже ее гибель. Эволюция человека пошла по пути межэкосистемного отбора вплоть до освоения всей биосферы. Поэтому исторический процесс с точки зрения биологии — слошная цепь массовых размножений людских популяций. Превентивных механизмов сохранения среды человечество не выработало, что с превращением его в глобальную силу грозит ему самоуничтожением. К этому вопросу мы вернемся в разделах 3.12 — 3.15. А сейчас продолжим обзор популяционных закономерностей в мире природы. Огромная экологическая литература подтверждает правило колебаний (цикличности) численности: никакая популяция не находится в состоянии абсолютной уравновешенности числа особей, обязательно помимо сезонных изменений численности особей возникают периодические флуктуации, обусловленные внешними по отношению к популяции факторами, и осциляции, связанные с собственными (внутренними) динамическими изменениями популяции. Эти вопросы столь подробно разработаны в широко доступной экологической литературе, что едва ли целесообразно их освещать здесь. Ограничимся констатацией упомянутого правила. Совершенно очевидно, что раз имеются ограничения согласно действию принципа минимального размера популяций и закона популяционного максимума, должно существовать обобщающее правило максимума размера колебаний плотности популяционного населения. Ю. Одум на с. 210 книги «Основы экологии» так формулирует это правило: «Существуют определенные верхние и нижние пределы для (средних — Н. Р.) размеров (речь идет о плотности населения — Н. Р.) популяции, которые соблюдаются в природе или которые теоретически могли бы существовать в течение сколь угодно длительного отрезка времени». При этом следует добавить: в условиях стабильности среды обитания. Ю. Одум совершенно справедливо замечает, что может быть 100 птиц на 1 га и 20 000 почвенных членистоногих на 1 м2, но никогда не бывает 20 000 птиц на 1 м2 и всего 100 членистоногих на 1 га. Правило максимума размера колебаний плотности популяционного населения может быть названо также законом количественной константности популяционного населения. Обсуждаемая закономерность указывает на среднесистемное число особей вида (его популяции) на единицу площади. Ясно, что отклонение от этого закона или правила (напомню, что слова «закон», «правило» и «принцип» употребляются как синонимы, исходя из удобства, а не каких-то других подходов) подчеркивает неблагополучие ситуации в регионе, служит биоиндикатором разлада в его экосистемах и шире — природных системах. В силу биотических и абиотических взаимосвязей отсутствие или даже лишь подавление численности какого-то вида неминуемо повлечет за собой цепь последствий, результат которых следует предвидеть. Если он отрицателен для людей, необходимо принять меры для нормализации обстановки. Для того чтобы закончить разбор закономерностей, регулирующих общую численность и плотность населения популяций, необходимо упомянуть правило максимальной рождаемости (воспроизводства): в популяции имеется тенденция к образованию теоретически максимально возможного количества новых особей. Оно достигается в идеальных условиях, когда отсутствуют лимитирующие экологические факторы, и размножение ограничено лишь физиологическими особенностями вида. Обычно же существует экологическая, или реализуемая рождаемость, возникающая в обычных или специфических условиях среды. Максимальная рождаемость есть константа, определяемая расчетным путем, например, умножением среднего числа гнезд, которые способна построить самка птицы за год на такое же число яиц, которые она может отложить в наиболее благоприятную часть сезона года. Максимальная рождаемость — тот предел, который характерен для скоростей увеличения числа особей в популяции. Очевидно, что правило максимальной рождаемости (воспроизводства) есть частный случай закона максимума биогенной энергии (энтропии) В. И. Вернадского — Э. С. Бауэра,(разд. 3.5.1). Стоит обратить внимание, что правило максимальной рождаемости (воспроизводства) действует в рамках закона (правила) популяционного максимума и принципа стабилизации экологической ниши (см. конец этого раздела). Очевидным следствием правила максимальной рождаемости (воспроизводства) популяции служит правило стабильности ее возрастной структуры: любая естественная популяция стремится к стабильной возрастной структуре, четкому количественному распределению особей по возрастам. Это правило сформулировано в 1925 г. А. Лоткой. Совершенно очевидно, что правило А. Лотки приложимо лишь к высшим организмам с возрастной структурой популяций и не имеет свойств универсальности для всех систематических групп, хотя в более широком биосистемном смысле оно универсально. Для многих организмов правило стабильности возрастной структуры популяции следует дополнить правилом стабильности соотношения полов (если дифференциация по полу вообще существует, что бывает отнюдь не всегда). В совокупности эти два правила составляют общее правило стабильности половозрастной структуры популяции. Перейдем к анализу закономерностей территориальной «микроструктуры» популяций, оставляя макроструктурные, биогеографические обобщения для следующего раздела 3.7. Естественно начать этот обзор с принципа территориальности: все особи и функциональные их объединения в популяции обладают индивидуальным и групповым пространством, возникающим в результате действия механизмов активного разобщения — поведенческого у высших организмов или физико-химического (аллелопатия и др.) у низших. Отличают индивидуальные и групповые (гнездовые, семейные) участки обитания. Примеры проявления территориальности можно найти в любом учебнике экологии. Столь же хорошо известен принцип скопления (агрегации) особей В. Олли, сформулированный в 1931 г.: скопление (агрегация) особей усиливает конкуренцию между индивидами, но способствует выживанию группы в целом; следовательно, как перенаселенность, так недонаселен-ность, препятствующая агрегации, могут служить лимитирующими факторами. Принцип агрегации особей диктует необходимость образования стай, стад, колоний, оптимальной густоты и распределения растений в посевах и имеет другие практические следствия*. Азбучной истиной в экологии служит деление типов распределения особей на скученное (небольшими группами), случайное и равномерное.
* Для социальной агрегации, образования человеческих общностей, а затем и поселений, достигающих в наши дни размера мегаполисов, существуют аналоговые закономерности, рассматриваемые в теории населенных мест, относящейся к экономической и социальной географии. Но первичный толчок и для человека был по происхождению биоэкологическим. Изначальная агрегация людей в группы с социализацией приобрела другие масштабы и стала развиваться по.особым путям. ** Реймерс Н. Ф., Воронов Г. А. Насекомоядные и грызуны верхней Лены. Иркутск: 1963. 191 с.
В 1950-х гг. я, видимо, впервые широко применил понятие кружева ареала и использовал его в практической работе при изучении мелких млекопитающих верхней Лены** (рис. 3.3). Книга, изданная в Иркутске тиражом в 500 экземпляров, осталась мало известной. Само понятие кружева ареала как неравномерности топографического распределения и плотности населения особей популяции с пространствами, где особи вида встречаются постоянно, редко, изредка и нерегулярно и вовсе не живут, кажется, вошло в научный обиход (рис. 3.4). Еще в большей мере принято рассматривать географическую неравномерность распределения особей в пространстве ареала, но это уже скорее биогеографическая, а не экосистемно-популяционная закономерность. Правило топографического, или популяционного, кружева ареала заключается в том, что популяция заселяет пространство неравномерно, оставляя «пустые» места, которые непригодны для ее жизни, и распадаясь на экологически разнородные микропопуляции, каждая из которых приурочена к определенному местообитанию. Связано это со многими факторами среды, как абиотическими, так и биотическими (в том числе антропогенными). Например, в долинах рек юга Средней Сибири, где имеются сельхозугодья, лесные полевки регулярно собираются на зимовку в стогах сена и скирдах соломы, что они не могут делать на таежных водоразделах, где таких стогов нет и никогда не было. Выше было упомянуто, что закон (правило) популяционного максимума и правило пищевой корреляции имеют более общий и глубокий экологический смысл. При этом была сделана ссылка на принцип стабилизации экологических ниш, или принцип биоценотической коэволюции: эволюция популяций внутри сообщества и эволюция этого сообщества скоррелированы таким образом, что каждый вид устойчиво сохраняет в этой природной системе свое функциональное место (экологическую нишу) до тех пор, пока внешние силы (воздействия надсистем или других аналогичных систем) не изменят существующего баланса. В данном случае слово «эволюция» понимается не только в историческом и надисторическом интервале времени, но и как динамическое понятие, т. е. как процесс постоянного взаимодействия видов, составляющих биотическое сообщество. Рост плотности населения популяции, заполнение ею емкости местообитания, в том числе потребление пищевых ресурсов, превентивно ограничено необходимостью остаться в составе материнского сообщества, не разрушить его как среду обитания рассматриваемой популяции в любых интервалах времени. Если происходит массовое размножение популяций какого-товида, то оно постепенно или относительно мгновенно гасится биоценоти-ческими механизмами (болезнями, паразитами, хищниками и т. п.). Коль этого не происходит (например, при акклиматизации и последующем массовом размножении нового для сообщества вида, не имеющего адекватных механизмов ограничения численности), меняется вся экосистема, и в конце концов либо возникает новый баланс, либо внедрившийся вид бывает элиминирован. Иногда он остается в составе сообщества как «равноправный» его участник, обычно немногочисленный. Это означает, что в сообществе был функциональный резерв («свободная» экологическая ниша), или сам вид выработал такие свойства (стабилизировал свою экологическую нишу), позволившие ему стать членом нового для него сообщества. В любом случае сообщество функционально изменилось, приобрело новые свойства, возможно, физиономически незаметные, но существенные для корреляции взаимоотношений между видами в нем.
Рис. 3.3. Картосхема основных биотопов, занимаемых элементарными фаунистическими комплексами территории верхнеленского ключевого участка. A. Биотопы фаунистических комплексов высоких водоразделов и расчленяющих их верховий речек и ручьев: 1 — биотоп фаунистического комплекса темнохвойного (лиственнично-елово-пихтово-кедрового) леса; 2 — биотоп фаунистического комплекса темнохвойного леса с участием лиственных пород (березы и осины); 3 — биотоп фаунистического комплекса гарей на месте темнохвойного леса, поросших осиново-березовым лесом различного возраста; 4 — биотоп фаунистического комплекса приручьевых участков темнохвойного (лиственнично-пихтово-елово-кедрового) леса; 5 — биотоп фаунистического комплекса приручьевых участков темнохвойного леса с участием лиственных пород (березы); 6 — биотоп фаунистического комплекса гарей на месте приручейных участков темнохвойного леса; 7 — биотоп фаунистического комплекса светлохвойных (лиственничных, сосново-лиственничных и лиственнично-сосновых) лесов. Б. Биотопы фаунистических комплексов коренных берегов долин крупных и средних рек с прилегающими к ним низкими водоразделами: 8 — биотоп фаунистического комплекса островных участков темнохвойных лесов с участием лиственных пород (березы, реже осины) среди светлохвойных массивов; 9 — биотоп фаунистического комплекса гарей на месте островных участков темнохвойных лесов (среди светлохвойных массивов), поросших осиново-березовым лесом различного возраста; 10 — биотоп фаунистического комплекса светло-хвойных (лиственнично-осиновых и сосново-лиственничных) лесов; 11 — биотоп фаунистического комплекса светлохвойных лесов с участием лиственных пород (березы и осины); 12 — биотоп фаунистического комплекса гарей на месте светлохвойных лесов, поросших березово-осиновым лесом различного возраста; 13 — биотоп фаунистического комплекса остепненных светлохвойных (лиственнично-сосновых и сосновых) лесов; 13а — биотоп фаунистического комплекса гарей на месте остепненного светлохвойного леса, поросшего лиственнично-березово-сосновыми молодняками и кустарниками; 14 — биотоп фаунистического комплекса безлесных гарей на месте остепненных светлохвойных лесов и остепненных луговых склонов («степоиды»). B. Биотопы фаунистических комплексов долин крупных и средних рек с низовьями впадающих в них других рек и ручьев: 15 — биотоп фаунистического комплекса приречных ельников с участием лиственных пород (березы, тополя) и небольших островов приречных ельников среди лугов поймы; 16 — биотоп фаунистического комплекса ивовых и черемуховых зарослей в поймах рек и ручьев; 17 — биотоп фаунистического комплекса суходольных и заливных пойменных лугов; 18 — биотоп фаунистических комплексов приручейных ельников с участием лиственных пород (березы); 19 — биотоп фаунистического комплекса гарей на месте приречных ельников, поросших березняком различного возраста; 20 — биотоп фаунистического комплекса ерниковых зарослей. Г. Биотопы фаунистических комплексов полей, огородов и населенных пунктов: 21 — биотоп фаунистических комплексов возделываемых полей; 22 — биотоп фаунистических комплексов открытых залежей и залежей с сосновой и елово-сосновой порослью 5 — 7-летнего возраста; 23 — биотоп фаунистических комплексов населенных пунктов (огородов и зданий). (Реймерс Н. Ф., Воронов Г. А., 1963)
Принцип стабилизации экологических ниш весьма существенен в практике акклиматизационных работ. Как правило, «свободных» экологических ниш, не возникших из-за нарушения экосистем человеком, быть не может. Любое сообщество находится в состоянии баланса, достигнутого в ходе биоценотической коэволюции: деятельность комплекса видов продуцентов управляется консументами и вводится в обратную связь редуцентами. Если бы такого баланса не существовало, экосистема деградировала бы. И поскольку этого не происходит, разговоры о «свободных» экологических нишах не имеют под собой никакого основания. Расхожая теоретическая ошибка о якобы имеющихся «свободных» экологических нишах приводит и уже многократно приводила к нелепым и к тому же весьма дорогостоящим акциям по акклиматизации животных и растений. Они в подавляющем большинстве случаев кончались неудачей, а иногда и приносили вред. Вопреки стабилизации экологических ниш вновь возникшие внутри популяции генотипы могут образовывать экологические субииши, расширять использование ресурсов среды. Их прежний экологический статус при этом не имеет значения: ранее они могли быть слабыми конкурентами. Это положение известно как принцип, или теорема В. Людвига, сформулированная в 1950 г. Теорема В. Людвига очень важна для понимания действия правила обязательности заполнения экологических ниш (разд. 3.8.2). Свод теоретических обобщений, часть из которых перекрывает друг друга, закончим упоминанием теории, или принципа, А. Николсона (1933): популяции суть стабильные системы, способные противостоять факторам внешней среды и контролировать эти факторы изменением своей плотности населения. Скорее можно говорить не о контроле среды, а о компенсациях в связи с ее изменением путем колебаний численности популяци-онного населения. Изменяя среду жизни популяций, человек должен ожидать компенсационных процессов. Не всегда они в его. интересах. Что касается относительной стабильности популяций как систем, то это свойство возвращает нас к правилу объединения в популяции, с которого начат этот раздел.
Рис. 3.4. Характер размещения и плотности населения азиатских лесных мышей на территории верхнеленского ключевого участка (окрестности дер. Якимовка Жигаловского района Иркутской области): 1 — балл 0 численности — зверьки не отловлены, они здесь не размножаются и редко забегают; 2 — балл 1 численности — встречаются единичные особи; 3 — балл 2 численности — зверьки постоянно живут и размножаются в биотопе, и численность вида довольно значительна; 4 — балл 3 численности — максимальная населенность биотопа данным видом. (Рис. 13, легенда рис. 5— стр. 84 и 43 книги «Насекомоядные и грызуны в верхней Лены»)
Вместе с группой закономерностей адаптации биосистем (разд. 3.4.2), действующих также в основном на популяционном уровне, изложенные здесь постулаты составляют костяк руководящих теоретических положений, объясняющих многие особенности функционирования популяционных совокупностей особей в рамках сообщества, т. е. относительно небольших пространств. В следующем разделе сделана попытка анализа территориально более широких биогеографических закономерностей. Конечно, такое разделение «по пространству» весьма условно. Любое функциональное взаимодействие происходит в рамках какой-то территории, а пространственная закономерность всегда функциональна.
studopedia.su - Студопедия (2013 - 2024) год. Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав!Последнее добавление
 Рис. 3.3. Картосхема основных биотопов, занимаемых элементарными фаунистическими комплексами территории верхнеленского ключевого участка. A. Биотопы фаунистических комплексов высоких водоразделов и расчленяющих их верховий речек и ручьев: 1 — биотоп фаунистического комплекса темнохвойного (лиственнично-елово-пихтово-кедрового) леса; 2 — биотоп фаунистического комплекса темнохвойного леса с участием лиственных пород (березы и осины); 3 — биотоп фаунистического комплекса гарей на месте темнохвойного леса, поросших осиново-березовым лесом различного возраста; 4 — биотоп фаунистического комплекса приручьевых участков темнохвойного (лиственнично-пихтово-елово-кедрового) леса; 5 — биотоп фаунистического комплекса приручьевых участков темнохвойного леса с участием лиственных пород (березы); 6 — биотоп фаунистического комплекса гарей на месте приручейных участков темнохвойного леса; 7 — биотоп фаунистического комплекса светлохвойных (лиственничных, сосново-лиственничных и лиственнично-сосновых) лесов. Б. Биотопы фаунистических комплексов коренных берегов долин крупных и средних рек с прилегающими к ним низкими водоразделами: 8 — биотоп фаунистического комплекса островных участков темнохвойных лесов с участием лиственных пород (березы, реже осины) среди светлохвойных массивов; 9 — биотоп фаунистического комплекса гарей на месте островных участков темнохвойных лесов (среди светлохвойных массивов), поросших осиново-березовым лесом различного возраста; 10 — биотоп фаунистического комплекса светло-хвойных (лиственнично-осиновых и сосново-лиственничных) лесов; 11 — биотоп фаунистического комплекса светлохвойных лесов с участием лиственных пород (березы и осины); 12 — биотоп фаунистического комплекса гарей на месте светлохвойных лесов, поросших березово-осиновым лесом различного возраста; 13 — биотоп фаунистического комплекса остепненных светлохвойных (лиственнично-сосновых и сосновых) лесов; 13а — биотоп фаунистического комплекса гарей на месте остепненного светлохвойного леса, поросшего лиственнично-березово-сосновыми молодняками и кустарниками; 14 — биотоп фаунистического комплекса безлесных гарей на месте остепненных светлохвойных лесов и остепненных луговых склонов («степоиды»). B. Биотопы фаунистических комплексов долин крупных и средних рек с низовьями впадающих в них других рек и ручьев: 15 — биотоп фаунистического комплекса приречных ельников с участием лиственных пород (березы, тополя) и небольших островов приречных ельников среди лугов поймы; 16 — биотоп фаунистического комплекса ивовых и черемуховых зарослей в поймах рек и ручьев; 17 — биотоп фаунистического комплекса суходольных и заливных пойменных лугов; 18 — биотоп фаунистических комплексов приручейных ельников с участием лиственных пород (березы); 19 — биотоп фаунистического комплекса гарей на месте приречных ельников, поросших березняком различного возраста; 20 — биотоп фаунистического комплекса ерниковых зарослей. Г. Биотопы фаунистических комплексов полей, огородов и населенных пунктов: 21 — биотоп фаунистических комплексов возделываемых полей; 22 — биотоп фаунистических комплексов открытых залежей и залежей с сосновой и елово-сосновой порослью 5 — 7-летнего возраста; 23 — биотоп фаунистических комплексов населенных пунктов (огородов и зданий). (Реймерс Н. Ф., Воронов Г. А., 1963)
Рис. 3.3. Картосхема основных биотопов, занимаемых элементарными фаунистическими комплексами территории верхнеленского ключевого участка. A. Биотопы фаунистических комплексов высоких водоразделов и расчленяющих их верховий речек и ручьев: 1 — биотоп фаунистического комплекса темнохвойного (лиственнично-елово-пихтово-кедрового) леса; 2 — биотоп фаунистического комплекса темнохвойного леса с участием лиственных пород (березы и осины); 3 — биотоп фаунистического комплекса гарей на месте темнохвойного леса, поросших осиново-березовым лесом различного возраста; 4 — биотоп фаунистического комплекса приручьевых участков темнохвойного (лиственнично-пихтово-елово-кедрового) леса; 5 — биотоп фаунистического комплекса приручьевых участков темнохвойного леса с участием лиственных пород (березы); 6 — биотоп фаунистического комплекса гарей на месте приручейных участков темнохвойного леса; 7 — биотоп фаунистического комплекса светлохвойных (лиственничных, сосново-лиственничных и лиственнично-сосновых) лесов. Б. Биотопы фаунистических комплексов коренных берегов долин крупных и средних рек с прилегающими к ним низкими водоразделами: 8 — биотоп фаунистического комплекса островных участков темнохвойных лесов с участием лиственных пород (березы, реже осины) среди светлохвойных массивов; 9 — биотоп фаунистического комплекса гарей на месте островных участков темнохвойных лесов (среди светлохвойных массивов), поросших осиново-березовым лесом различного возраста; 10 — биотоп фаунистического комплекса светло-хвойных (лиственнично-осиновых и сосново-лиственничных) лесов; 11 — биотоп фаунистического комплекса светлохвойных лесов с участием лиственных пород (березы и осины); 12 — биотоп фаунистического комплекса гарей на месте светлохвойных лесов, поросших березово-осиновым лесом различного возраста; 13 — биотоп фаунистического комплекса остепненных светлохвойных (лиственнично-сосновых и сосновых) лесов; 13а — биотоп фаунистического комплекса гарей на месте остепненного светлохвойного леса, поросшего лиственнично-березово-сосновыми молодняками и кустарниками; 14 — биотоп фаунистического комплекса безлесных гарей на месте остепненных светлохвойных лесов и остепненных луговых склонов («степоиды»). B. Биотопы фаунистических комплексов долин крупных и средних рек с низовьями впадающих в них других рек и ручьев: 15 — биотоп фаунистического комплекса приречных ельников с участием лиственных пород (березы, тополя) и небольших островов приречных ельников среди лугов поймы; 16 — биотоп фаунистического комплекса ивовых и черемуховых зарослей в поймах рек и ручьев; 17 — биотоп фаунистического комплекса суходольных и заливных пойменных лугов; 18 — биотоп фаунистических комплексов приручейных ельников с участием лиственных пород (березы); 19 — биотоп фаунистического комплекса гарей на месте приречных ельников, поросших березняком различного возраста; 20 — биотоп фаунистического комплекса ерниковых зарослей. Г. Биотопы фаунистических комплексов полей, огородов и населенных пунктов: 21 — биотоп фаунистических комплексов возделываемых полей; 22 — биотоп фаунистических комплексов открытых залежей и залежей с сосновой и елово-сосновой порослью 5 — 7-летнего возраста; 23 — биотоп фаунистических комплексов населенных пунктов (огородов и зданий). (Реймерс Н. Ф., Воронов Г. А., 1963)
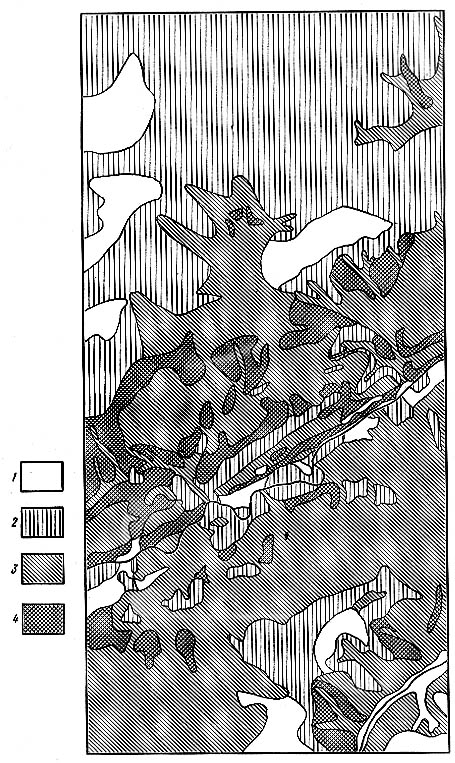 Рис. 3.4. Характер размещения и плотности населения азиатских лесных мышей на территории верхнеленского ключевого участка (окрестности дер. Якимовка Жигаловского района Иркутской области): 1 — балл 0 численности — зверьки не отловлены, они здесь не размножаются и редко забегают; 2 — балл 1 численности — встречаются единичные особи; 3 — балл 2 численности — зверьки постоянно живут и размножаются в биотопе, и численность вида довольно значительна; 4 — балл 3 численности — максимальная населенность биотопа данным видом. (Рис. 13, легенда рис. 5— стр. 84 и 43 книги «Насекомоядные и грызуны в верхней Лены»)
Рис. 3.4. Характер размещения и плотности населения азиатских лесных мышей на территории верхнеленского ключевого участка (окрестности дер. Якимовка Жигаловского района Иркутской области): 1 — балл 0 численности — зверьки не отловлены, они здесь не размножаются и редко забегают; 2 — балл 1 численности — встречаются единичные особи; 3 — балл 2 численности — зверьки постоянно живут и размножаются в биотопе, и численность вида довольно значительна; 4 — балл 3 численности — максимальная населенность биотопа данным видом. (Рис. 13, легенда рис. 5— стр. 84 и 43 книги «Насекомоядные и грызуны в верхней Лены»)