КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Пол у Е. соli
|
|
|
|
В 1946 году Джошуа Ледерберг и Эдвард Тэйтем принялись ставить генетические эксперименты на бактериях. Несколькими годами ранее Тэйтем работал в сотрудничестве с Джорджем Биллом, и они на основе опытов с плесенью Neurospom выдвинули теорию «один ген — один фермент», оказавшую столь большое влияние на развитие генетики. Ледерберг и Тэйтем надеялись при помощи более простых организмов сделать еще более грандиозное открытие. Лурия и Дельбрюк к тому времени уже доказали, что бактерии подобно другим организмам могут мутировать и образовывать разные штаммы, полезные в исследованиях. Однако для генетических экспериментов требуется скрещивать различные линии организмов с половым размножением, тогда как у бактерий, как считали в то время, пола нет. Ледерберг и Тэйтем предположили, что если бы у бактерий имелся пол, то разные клетки с разным генотипом скрещивались бы между собой и давали рекомбинантов, как и более высокоразвитые организмы. Поэтому если обнаружить рекомбинантные организмы, то это будет доказательством того, что у бактерий имеется нечто вроде полового размножения. Они предположили, что даже в этом случае количество рекомбинантов будет очень малым, и поэтому нужно постараться вывести рекомбинантов между двумя ауксотрофными штаммами. Один из штаммов для своего роста требует треонин и лейцин, но может производить свои метионин и тиамин (генотип thr--leu--met+hti); другой штамм может вырабатывать треонин и лейцин, но требует метионин и тиамин (генотип thr+leu+met~thi~). При смешении этих штаммов действительно обнаруживались прототрофы, которые могли образоваться только в результате рекомбинации.
Леденберг обнаружил, что рекомбинации могут происходить только между некоторыми фертильными штаммами, которые он обозначил F+, и другими штаммами с довольно низкой частотой, которые он обозначил F--, притом с довольно низкой частотой. Фертильность у бактерий — это нечто вроде инфекции, потому что когда клетки F--смешиваются с клетками F+, они преобразуются в клетки F+. Такое странное явление прояснили эксперименты Уиляма Хэйза из Великобритании, Франсуа Жакоба и Эли Вуллмана из Франции и Л. Л. Кавалли из Италии. Сначала они открыли, что клетки F+ содержат генетический фактор, названный ими F-фактором, или фертильным фактором. Как мы увидим далее, F-фактор представляет собой отдельную молекулу ДНК. При контакте с другими клетками клетки F+ способны передавать копии своих F-фак-торов другим клеткам, преобразуя их в клетки типа F+. С низкой вероятностью они могут заодно передавать и некоторые свои гены клеткам F--, и так происходят рекомбинации. Затем Хэйз и Кавалли открыли вариант штамма F+, названный Hfr (high frequency of recombination — «высокая частота рекомбинации»), который передает свои гены клеткам F-- с довольно высокой частотой. Механизм рекомбинации стал ясен после скрещиваний прототрофного штамма Hfr, чувствительного к стрептомицину, со множественным ауксот-рофным штаммом F--, устойчивым к антибиотику, и с генотипом strrthr~leu~met~lac~gal~thi~. Каждый ген означает метаболический недостаток, например, неспособность синтезировать треонин {thr-) или неспособность расти на лактозе (lac-). При смешивании клетки Hfr и F- спариваются в процессе конъюгации (слияния). Через некоторое время их помещают в среду со стрептомицином, чтобы убить клетки Hfr и проверить, какие дикие гены клетки F- унаследовали от клеток Hfr. В ставшем классическом эксперименте Жакоб и Вуллман через разные промежутки времени брали образцы конъюгирующих клеток и разделяли их в смесителе, после чего выращивали в различных средах и определяли унаследованные гены. Этот эксперимент показал, что каждый маркер Hfr начинает проявляться через строго определенный промежуток времени: ген В только через семь минут, ген С — через девять, ген D — только через 15 минут и т. д. Отсюда следует, что Hfr передает клеткам F- гены в линейной последовательности, то есть в той последовательности, в которой они соединены в хромосоме бактерий.
На рис. 10.1 объясняются полученные результаты. Фактор F в клетках F+ представляет собой небольшую кольцевую молекулу ДНК. Контакт с клетками F- побуждает ее к быстрой репликации и переносу реплики в клетки F-, благодаря чему они превращаются в клетки F+. Во время этого процесса некоторые гены F+ случайным образом проникают в клетку F--, хотя до сих пор не совсем ясно, как это происходит. Как бы то ни было, в клетках Hfr 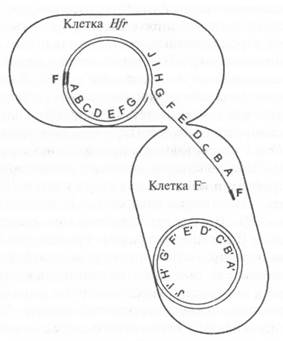
Рис. 10.1. Перенос ДНК при скрещивании Hfr x F. Клетка Hfr соединяется с клеткой F--, и между ними образуется конъюгационная трубка. Фактор F, встроенный в хромосому Hfr, начинает реплицироваться. Но так как фактор F встроен в хромосому, он заодно запускает репликацию всей хромосомной ДНК. Поэтому в клетку F-- переходит копия хромосомы Hfr с генами в линейной последовательности. Как только донорская ДНК проникает в клетку F--, она может спариваться с хромосомой F-- и образовывать рекомбинанты с аллелями Hfr в хромосоме F--
фактор F встраивается в хромосому бактерии, которая тоже кольцевая, хотя и больше по размеру. Теперь при контакте с F-- фактор F снова начинает переносить свою копию, но поскольку фактор F и бактериальная хромосома представляют собой единое целое, эта копия содержит бактериальные гены, которые передаются последовательно: А, В, С, D и т. д. Конъюгация редко длится так долго (90—100 минут), чтобы произошла репликация всей кольцевой хромосомы вплоть до начала фактора F, поэтому клетки F--, как правило, не так уж часто преобразуются в клетки Hfr.
После конъюгации вновь перенесенная ДНК Hfr рекомбинирует с хромосомой F--, и появляются разнообразные рекомбинанты. Но в данном случае по частоте рекомбинации определяют не общую карту Е. coli и расстояние между генами в единицах, а расстояние в минутах между маркерами. Фактор F может встраиваться в различные места и переноситься в разных направлениях от этой точки; каждое такое встраивание приводит к образованию отдельного штамма Hfr уникального происхождения. Так строят карту различных участков генома. Подробные генетические карты Е. coli (рис. 10.2) и других бактерий были составлены при исследовании всех этих штаммов, а также при помощи других методов, которые будут описаны далее.
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 474; Нарушение авторских прав?; Мы поможем в написании вашей работы!