КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Розділ 4. Аналіз причин порушення менделівських закономірностей розщеплення при моногенному успадкуванні ознак
|
|
|
|
У природі досить часто відхилення від ідеальних менделівських співвідношень фенотипових класів при розщепленні проявляються систематично, а використання статистичних методів аналіза лише доводить їх невипадковість. Для того, щоб розібратися у причинах таких відхилень, необхідно згадати, при яких умовах завжди проявляються визначені Г.Менделем статистичні закономірності розщеплення у гібридному потомстві.
Успадкування здійснюється у відповідності із менделевськими формулами розщеплення, якщо:
1) гени локалізуються в різних хромосомах або на досить значній відстані в одній хромосомі;
2) різні типи гамет формуються та виживають в однакових співвідношеннях (рівноймовірно);
3) генетично різні типи зигот і відповідні генотипи виникають і виживають з однаковою вірогідністю;
4) функція генів проявляється повністю, отже спостерігається повна експресивність і повна пенетрантність генів;
5) спостерігається повна домінантність ознаки;
6) досліди проводяться на великій вибірці;
7) проявлення ознаки не залежить від зовнішніх умов і від генотипу.
Найчастіше причинами порушень менделівського розщеплення за фенотипом при моногенному успадкуванні є:
1) статистичні причини;
2) неповне домінування та наддомінування генів;
3) множинний алелізм і кодомінування;
4) диференційна смертність особин – носіїв певних генотипів;
5) неповне проявлення функції гена при певних умовах.
Відомо, що розщеплення при схрещуваннях мають імовірнісний характер, тому практично завжди у дослідах спостерігаються відхилення від теоретично очікуваного розщеплення. Ці відхилення є випадковими та в значній мірі залежать від величини вибірки. Крім того, відхилення можуть бути наслідком порушення будь-якої з умов менделівського успадкування (загибель гамет або зигот певного генотипу, зчеплення генів тощо). Тому при аналізі розщеплення для розуміння причини відхилення спочатку необхідно оцінити їх величину та значущість. З цією метою найчастіше використовують метод χ2. Сутність методу полягає у зіставленні теоретично розрахованого розщеплення на основі нульової гіпотези (Но) з даними досліду за всіма класами розщеплення за формулою c2 (хі – квадрат): 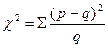 ;
;
|
|
|
де р – фактична кількість особин у кожному фенотиповому класі;
q – теоретично розрахована кількість особин у цьому класі за законами менделівського успадкування,
S - сума показників всіх класів.
Результати досліду (р) та теоретичне очікування (q) заносяться в таблицю. Надалі визначають величину відхилення (d), яку возводять в квадрат (d2) для унеможливлення появи негативних величин. Квадрат відхилення ділять на теоретично очікувану для кожного класа величину (d2: q). Сума цих величин є показником χ2. Після цього за таблицею Фішера з урахуванням ступенів свободи визначають ймовірність випадковості відхилення.
Кількість ступенів свободи розраховується за формулою:
n’= n – 1, де n – кількість фенотипових класів розщеплення в гібридному потомстві.
Таблиця значень c2 при різних ступенях свободи
(за Фішером, із скороченням)
| Кількість ступенів свободи (nI) | Імовірність випадковості відхилення, р | |
| 0,05 | 0,01 | |
| 3,841 | 6,635 | |
| 5,991 | 9,210 | |
| 7,815 | 11,341 | |
| 9,488 | 13,277 | |
| 11,070 | 15,086 | |
| 12,592 | 16,812 | |
| 14,067 | 18,475 |
Значення χ2, наведені в таблиці, вказують межі, в яких дослідні результати відповідають теоретично очікуваним. При повній відповідності дослідних і теоретичних даних χ2 дорівнює 0. Якщо χ2 ≠0, то різниці величин, що порівнюються, є випадковими (нульова гіпотеза). Ймовірність, яка вказана у таблиці – це ймовірність цієї нульової гіпотези. Допустимою межею імовірності у статистиці прийнято вважати величину 0,05. Наприклад, ймовірність 0,05 говорить: якщо величини, які порівнюються, відрізняються випадково, то табличне значення χ2 може проявитися тільки у 5 вибірках зі 100 подібних. У статистиці прийнято вважати, що події, які відбуваються з ймовірністю 0,05 і менше, практично не зустрічаються. Нульова гипотеза відкидається, якщо значення χ2 є більшим за те, що наведено в стовбці р = 0,05 при відповідній кількості ступенів свободи.
|
|
|
Отже, якщо значення χ2 перевищують табличні при відповідних ступенях свободи, то це означає, що відхилення у досліді не є випадковим, воно не може бути пояснено причинами статистичного характеру. Тому при значенні c2, рівному або більшому за значення, вказане у таблиці, вважають різниці величин, що порівнюються, суттєвими та закономірними.
У решті випадків, коли χ2 < табличного значення, відмінності між теоретично розрахованими і фактично одержаними результатами експерименту вважають випадковими, у межах допустимої похибки.
Таким чином, метод χ2 дозволяє встановити відповідність дослідних та очікуваних результатів за кожним класом та за всім розщепленням у цілому. Слід пам’ятати, що цей метод не використовується для значень, виражених у відносних числах та у процентах, а також для вибірок з кількістю особин у будь-яких теоретичних класах меньше п’яти. На основі критерію χ2 відкидають помилкові гіпотези, якщо відхилення є значущим, коли не існує відповідності між дослідними та теоретично розрахованими величинами.
У випадку неповного домінування (АА > Аа) розщеплення в F2 за фенотипом співпадає з розщепленням за генотипом 1:2:1 (1/4 АА: 2/4 Аа: ¼ аа), Це відбувається, коли домінантний алель відповідає за синтез активної форми білка – фермента, а рецесивний алель контролює той самий білок, але із зниженою ферментативною активністю. Це явище реалізується у гетерозигот у вигляді неповного домінування, при якому гібрид F1 має свій власний фенотип, відмінний від фенотипу батьків. Прояв ознак у гетерозигот є проміжним, із більшим або меншим відхиленням від домінантного та рецесивного станів гена. Отже, за проміжного успадкування всі особини F1 є фенотипово одноманітними, але в F2 розщеплення за фенотипом співпадає з розщепленням за генотипом.
|
|
|
При наддомінуванні домінантний алель у гетерозиготному стані має більш сильне проявлення, ніж у гомозиготному: АА<Aa>aa. Це явище лежить в основі проявлення ефекту гетерозису.
В експериментах Грегора Менделя (1822 – 1886) кожний ген представлений лише двома алелями (домінантний і рецесивний). Але більшість (а можливо і всі) гени у популяціях організмів представлені декількома алелями (множинні алелі), отже, існують більш ніж в двох алельних станах, які виникають унаслідок генних мутацій. Це явище називають множинним алелізмом.
Множинні алелі виникають у результаті багаторазового мутування того ж самого локусу в хромосомі. Окрім основних (домінантного та рецесивного) алелів гена з’являються проміжні алелі, котрі по відношенню до домінантного поводять себе як рецесивні, а по відношенню до рецесивного – як домінантні алелі того ж самого гена. Взаємодія продуктів активації множинних алелів фенотипово проявляється у вигляді міжалельної комплементації.
Міжалельна комплементація спостерігається у компаундів (а1 а2,, а2, а3, і т.д.), якщо продуктом мутантного гена (а) слугує поліпептид, що є субодиницею білка – гомомультимера. Об’єднання у компаунд двох рецесивних алельних генів, кожний з яких кодує мутантний поліпептидний ланцюг, призводить до відтворення ознаки дикого типу, тобто із двох типів частково ушкоджених субодиниць збирається білок з майже нормальною функцією. У цих випадках рецесивні алелі того ж самого гена кодують поліпептиди з ушкодженнями різних доменів (певних функціональних ділянок) у молекулах. Якщо збірка четвертинного білка у компаунда здійснюється із таких субодиниць, здатних компенсувати функціональну активність одна одної, ферментативна активність цього білка буде відновлена. Тому схрещування двох фенотипово однакових мутантів призводить до утворення гібридів F1 дикого типу: а1а1 х а 2 а 2 = а1 а2. Міжалельна комплементація дуже нагадує результат комплементарної взаємодії генів, однак в останньому випадку взаємодіють продукти функціонування неалельних генів.
|
|
|
Відома велика кількість прикладів множинного алелізму.Три алелі одного гена визначають успадкування груп крові системи АВО, відкритих Карлом Ландштейнером (1868 – 1943) у 1900 році. У системі груп крові АВ0 на поверхні еритроцитів формуються два антигени під контролем генних алелів (ІА та ІВ) та існує третій алель, що не контролює синтез антигена. Але у кожної людини у диплоїдному генотипі можливе існування лише 2-х алелів цієї серії. Два антигени АВ0 є ізоантигенами і виявлені у більшості людей в еритроцитах, нормальних і пухлинних клітинах інших тканин, у мітохондріях і мікросомах, у біологічних рідинах організму – слині, шлунковому соку, молоці. Антигени А і В виявляють захисну дію на слизову оболонку шлунка та 12-палої кишки. Тому у людей з групою 0, у котрих немає типового антигена, часто розвивається виразкова хвороба шлунка і 12-палої кишки.
При взаємодії антигену з антитілами відбувається аглютинація – злипання еритроцитів, що призводить до закупорки судин і смерті людини. Якщо у геномі існують алелі ІА та ІВ, тоді синтезуються антигени А та В, при цьому в організмі не утворюються антитіла α та β. Якщо алелі А або В відсутні, тоді виникають антитіла α проти А та β проти антигена В.
Групи крові, що містять антиген А, є неоднорідними. У цій групі розрізняють три підгрупи – А1, А2, А3. Кожний з генів, що контролює ці антигени, може займати той самий локус хромосоми. Алель А1 > A2 > A3. Антиген А3 зустрічається дуже рідко, А1 складає ¾, А2 – ¼ всієї популяції.
Вважають, що еволюційно найдавнішою є група крові АВ (нині не існує). Аглютиногени (антигени) виникли еволюційно раніше, ніж аглютиніни (антитіла α та β). Пізніше, в результаті мутації виникла група крові 0. Найдавніша група крові (АВ) зникла у ході еволюції. Нинішня група АВ походить від людей з генотипами А і В.
При переливанні крові необхідно знати групи крові донора та реципієнта, щоб запобігти аглютинації (склеювання) еритроцитів донорської крові у кровотоку реципієнта. У системі АВО є чотири групи крові: О, А, В, АВ. Вони визначаються трьома алелями: ІА, ІВ, іо. Алелі ІА та ІВ є домінантними відносно алеля іо, але кодомінантними у відношенні один до одного (табл. 2.).
Таблиця 2.- Групи крові системи АВО
| Генотип | Фенотип (група крові) |
| ІАІА, ІАіо ІВІВ,ІВіо ІАІВ іоіо | А В АВ О |
Кодомінування – проявлення у гетерозиготному стані ознак, що детермінуються обома алелями. Наприклад, кожний з алельних генів кодує певний білок, а у гетерозиготного організму синтезуються вони обидва. У таких випадках шляхом біохімічного дослідження можна встановити гетерозиготність без проведення аналізуючого схрещування. За типом кодомінування у людини успадковується четверта група крові (АВ).
Отже, алелі ІА та ІВ при сполученні їх в зиготі визначають проявлення нової ознаки – группи крові АВ (ІV). При існуванні трьох алелів можливе існування шести різних генотипів.
Названі алелі контролюють синтез білкових молекул, вбудованих у мембрану еритроцитів – аглютиногенів. Аглютиногену (антиген) еритроцитів відповідає аглютинін (антитіло) плазми крові. Відомі два аглютиногени А і В, два аглютиніни α і β. У людей з І (О) групою аглютиногени в еритроцитах відсутні, а в плазмі є обидва аглютиніни (α і β). У людей з ІІ (А) групою еритроцити несуть аглютиноген А, а в плазмі міститься аглютинін β. У людей ІІІ (В) групи еритроцити несуть аглютиноген В, плазма – аглютинін α. У ІУ (АВ) групі еритроцити мають обидва аглютиногени, аглютиніни у плазмі відсутні.
Особи з І (О) групою є універсальними донорами: їхню кров можна переливати особам з будь-якою групою крові, оскільки їх еритроцити не містять антигенів. Особи з ІУ (АВ) групою є універсальними реціпієнтами: ним можна переливати кров будь-якої групи, оскільки їхня плазма не містить аглютинінів. Ввважається, що під час переливання крові аглютиніни плазми донора розбавляються плазмою реципієнта, тому склеювання може відбуватися тільки внаслідок взаємодії антигенів еритроцитів донора з аглютинінами плазми реципієнта. Тому реціпієнту з І (О) групою крові можна переливати кров донора тільки тієї ж групи, реціпієнту з ІІ (А) групою – кров донора І (О) або ІІ (А) груп, реціпієнту з ІІІ (В) групою – кров донора І(О) або ІІІ (В) груп. Кров від донора ІІ (А) групи можна переливати реципієнту ІІ (А) та ІУ (АВ) груп, від донора ІІІ (В) групи – реципієнту ІІІ (В) та ІУ (АВ) груп, від донора ІУ (АВ) групи – тільки реципієнту ІУ групи (табл.3).
Таблиця 3.- Можливість переливання крові особам різних груп
| Донор Реципієнт | І (О) Генотип ioio | II (A) генотип IA | III (B) генотип IB- | IV (AB) генотип IAIB |
| І (О) | + | - | - | - |
| ІІ (А) | + | + | - | - |
| ІІІ (В) | + | - | + | - |
| ІУ (АВ) | + | + | + | + |
Визначення груп крові має велике значення не тільки для медицини, але і для судово-слідчої практики.
Кількість можливих генотипів у випадку множинного алелізму залежить від кількості алелів даного гена. Якщо ген представлений єдиним алелем А, то і генотип за цим алелем може бути лише один – АА. Якщо алелів два, А1 та А2, то можливі три генотипи: два гомозиготних, А1А1 та А2А2, і один гетерозиготний: А1А2. У загальному випадку, при існуванні у популяції n- ної кількостіалелів, можливе існування 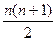 генотипів, з яких n гомозиготні, а решта
генотипів, з яких n гомозиготні, а решта 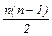 – гетерозиготні (табл. 4).
– гетерозиготні (табл. 4).
Таблиця 4. - Кількість можливих генотипів при n-ній кількості алелів
| Кількість алелів | Кількість генотипів | Кількість типів гомозигот | Кількість типів гетерозигот |
| N | n(n + 1)/2 | n | n(n - 1)/2 |
Відхилення, пов’язані із диференційною смертністю певних генотипів. У період становлення генетики були описані випадки, коли при схрещуванні особин з однаковим фенотипом постійно відбувалося розщеплення у співвідношенні 2:1. Нині відомі випадки такого розщеплення при схрещуванні жовтих за забарвленням мишей, платинових лисиць, сріблясто-соболиних норок, коротконогих курей, лінійних (луска тільки на боковій лінії) карпів, сірих каракулевих овець, людей, хворих на ахондроплазію (карлики з нормальними за розмірами головою і тулубом). Можна припустити, що у цьому випадку домінантний алель у гомозиготному стані є летальним. Дійсно, деякі гени у гомозиготному стані викликають загибель зиготи, тому відповідний клас особин в F2 не з’являється, хоча генотипи в F2 утворюються із звичайною ймовірністю (1АА:2Аа:1 аа), Гомозиготні організми за деякими (як рецесивними, так і домінантними) генами опиняються нежиттєздатними, Наприклад, домінантний алель сірого забарвлення шерсті каракульських овець у гомозиготному стані викликає загибель ягнят унаслідок недорозвинення травної системи. У людини аналогічно успадковується домінантний ген брахідактилії (короткопалості): ознака проявляється при гетерозиготності індивіда, у гомозигот цей ген призводить до загибелі зародків на ранніх етапах розвитку. У цих випадках мутантні гени є плейтропними, тобто здатними викликати не лише прояв певної ознаки, але й летальну дію (у гомозиготному стані).
Плейотропія – множинна дія гена, його здатність впливати одночасно на проявлення декількох ознак організму. Мутація одного гена призводить до порушення обміну на певному етапі метаболізму, але оскільки метаболічні шляхи у клітині є багатоетапними та взаємопов’язаними, то ця ланка порушеного обміну неминуче відображається на наступних етапах, отже, на проявленні деяких інших ознак. Тому розщеплення за фенотипом в F2 при моногібридному схрещуванні відбувається звичайно у співвідношенні 2/3 А-:1/3 аа (а не ¾ А-:1/4 аа), як передбачено менделівским законом розщеплення.
Відхилення, викликані неповним проявом функцій генів за даних умов, зустрічаються досить часто і можуть обумовлюватися неповною пенетрантністю або неповною експресивністю гена, що контролює ознаку. Обидва поняття – пенетрантність та експресивність – були введені у 1925 році М.В. Тимофєєвим-Ресовським для визначення варіюючого прояву гена, який супроводжується появою нових фенотипових класів залежно від умов навколишнього або генотипового середовища.
Пенетрантність гена – частота фенотипового проявлення гена в популяції особин, які є його носіями; виражається у процентах. Наприклад, пенетрантність гена 75% означає, що тільки у 75% особин, які мають в генотипі цей ген, проявляється його фенотиповий ефект.
Експресивність гена – сила дії гена, яка характеризується ступенем фенотипового проявлення ознаки, що контролюється цим геном. Експресивність залежить від взаємодії певного гена із зовнішніми умовами та генотиповим середовищем (дії інших генів).
Експресивність і пенетрантність ознаки залежать від особливостей генотипу, дози гена (кількості його копій у геномі), положення гена (місця його розташування на хромосомі).
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 1219; Нарушение авторских прав?; Мы поможем в написании вашей работы!