КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Строение синапса. Синаптическая передача возбуждения ….….. 23 3 страница
В некоторых межнейронных синапсах осуществляются параллельно и электрическая и химическая передача. В этих контактах синаптическая щель имеет участки со структурой электрического и химического синапсов. Такие синапсы получили название смешанных, или электрохимических.
Дегенеративные и регенеративные процессы в ЦНС
Дегенерация и гибель определённых нейронов, нервных волокон и синаптических окончаний являются неотъемлемыми составляющими процесса развития организма. Было обнаружено, что в течение короткого периода времени в самом начале эмбрионального развития, когда периферические нервные волокна устанавливают свои связи с иннервируемыми клетками-мишенями, происходит дегенерация большого числа нейронов в спинномозговых узлах и в моторных областях спинного мозга.
Гибель клеток в эмбриональный период – обычное явление для многих отделов нервной системы. Число гибнущих нейронов в некоторых случаях достигает 75 %. Часто отмечается совпадение момента гибели большого количества нейронов данного отдела ЦНС с моментом иннервации ими своих мишеней. Нейроны, аксоны которых достигают клеток-мишеней, выживают, т.к., по-видимому, получают от иннервируемых клеток какой-то сигнал или поддерживающий трофический фактор. Таким образом, при образовании синаптических контактов происходит конкуренция за мишени, укрепление нужных связей и устранение бесполезных и избыточных.
Дегенерируют не только нейроны, но и отдельные нервные окончания и синапсы. Например, начальный сегмент аксона у мотонейронов спинного мозга имеет синапсы, которые на более поздних стадиях эмбрионального развития исчезают. В мозжечке на ранней стадии развития тело клеток Пуркинье имеет много шипиков, которые образуют синапсы с лиановидными волокнами. Позже и эти шипики и соответствующие синапсы полностью исчезают. Предполагают, что ранние соединения помогают образованию других синапсов, или обеспечивают некоторый контроль над возбудимостью, который необходим на определённой стадии развития.
 После того как процессы развития полностью завершаются, новые нервные клетки не возникают, а если и возникают (обонятельные рецепторные клетки), то в незначительном числе. Однако, хотя новые нейроны не образуются, у каждого нейрона сохраняется способность к формированию новых отростков и новых синаптических контактов (рис. 17). Поэтому синаптические сети, образуемые между разными нейронами, подвергаются непрерывной модификации, особенно при овладении новыми навыками, познании и запоминании новой информации.
После того как процессы развития полностью завершаются, новые нервные клетки не возникают, а если и возникают (обонятельные рецепторные клетки), то в незначительном числе. Однако, хотя новые нейроны не образуются, у каждого нейрона сохраняется способность к формированию новых отростков и новых синаптических контактов (рис. 17). Поэтому синаптические сети, образуемые между разными нейронами, подвергаются непрерывной модификации, особенно при овладении новыми навыками, познании и запоминании новой информации.
Хотя нервная система не может порождать новые нейроны взамен утраченных, каждая клетка способна дать новые отростки взамен тех, которые были утрачены или повреждены. Также нервная система способна восстанавливать специфические синаптические связи. При отделении аксона от тела нейрона этот аксон дегенерирует. Однако вскоре на сохранившем связь с телом нейрона оставшемся кусочке аксона образуется колбообразный вырост. Он начинает медленно со скоростью 0,5-4,5 мм в сутки расти по тому же пути, который занимал дегенерировавший аксон. При нормальных условиях новый аксон дорастает до иннервируемого органа и образует синапсы с клетками-мишенями. Таким способом происходит регенерация отростков нейрона и восстанавливается нормальная иннервация органа или ткани.
Наряду с врождённой тенденцией к образованию новых связей при повреждении старых для нейронов также присуща способность устанавливать новые связи в случае утраты прежних постсинаптических мишеней. Потеря определённого типа синаптических окончаний стимулирует возникновение аксонных выростов (колб роста), обладающих подвижностью и способных прорастать к другим клеткам и устанавливать новые синаптические контакты. Эксперименты показывают, что если перерезается один вход к нейрону, то другой вход увеличивает количество своих нервных окончаний, занимая освободившиеся постсинаптические участки на теле и дендритах этого нейрона.
Предполагают, что любой нейрон запрограммирован на образование определённого количества синапсов и реагирует на повреждение нервной ткани таким образом, чтобы компенсировать потери и попытаться восстановить необходимое количество синапсов, не смотря даже на то, что они могут образоваться в несоответствующих местах. Если две нервные клетки должны взаимодействовать более интенсивно, то число связей между ними может возрастать за счёт образования новых синапсов при одновременном сохранении старых. В опытах установлено, что если у млекопитающего вырезать определённый участок мозга и пересадить его в мозг другого животного, то пересаженный участок даёт отростки, которые устанавливают связи с мозгом реципиента.
Синаптические медиаторы
В качестве синаптических медиаторов выступает не одно, а целая группа химически разнородных веществ. Среди медиаторов больше всего веществ с относительно небольшой молекулярной массой, однако и некоторые полипептиды выполняют роль передатчиков возбуждения в ряде синапсов центральной нервной системы. Медиатором считается вещество при условиях, если оно: 1) имеется в соответствующих пресинаптических окончаниях; 2) способно высвобождаться под влиянием нервного импульса; 3) способно изменять ионную проницаемость и электрический заряд постсинаптической мембраны; 4) по молекулярным и ионным механизмам действия на постсинаптическую мембрану действие вещества, высвобождаемого нервным импульсом, идентично действию этого же вещества, прикладываемого искусственно к постсинаптической мембране.
Из-за сложности строения мозга до сих пор точно не установлена химическая природа некоторых медиаторов в ЦНС. Обнаружено уже около 20 медиаторов и продолжается открытие новых. К ним относятся: ацетилхолин, норадреналин, дофамин, серотонин, гамма-аминомасляная кислота (ГАМК), глицин, вещество Р, АТФ, гистамин, соматостатин и др. Согласно принципу Дейла, каждый нейрон синтезирует и выделяет во всех своих синаптических окончаниях один и тот же медиатор. Нейроны, выделяющие ацетилхолин, называют холинергическими, норадреналин – адренергическими, серотонин – серотонинергическими.
Большому количеству различных медиаторов соответствует большое количество постсинаптических рецепторов, которые называют соответственно холинорецепторами, адренорецепторами, серотонинорецепторами, ГАМК-рецепторами и т.д. Этими мембранными рецепторами являются белковые молекулы, способные вступать во взаимодействие со специфическими медиаторами. В результате такого взаимодействия изменяется ионная проницаемость постсинаптической мембраны, что приводит к уменьшению или увеличению разности потенциалов между наружной и внутренней поверхностью мембраны, т.е. деполяризации или гиперполяризации.
Постсинаптические рецепторы обладают высокой чувствительностью к соответствующим медиаторам. Например, холинорецепторы реагируют на появление ацетилхолина в синаптической щели в концентрации 10-8 моль/л. Чрезмерно длительное действие медиатора на соответствующие рецепторы приводит к десенситизации рецепторов, т.е. к снижению чувствительности этих рецепторов к медиатору.
Один и тот же медиатор может вступать в реакцию с различными постсинаптическими рецепторами и вызывать противоположные эффекты. Например, ацетилхолин в нервно-мышечных синапсах скелетных мышц у человека действует на так называемые Н-холинорецепторы (чувствительные к никотину), которые открывают натриевые каналы в постсинаптической мембране, что вызывает деполяризацию и возникновение ВПСП. В синапсах, образованных окончаниями блуждающего нерва на клетках сердца этот же ацетилхолин действует на М-холинорецепторы (чувствительные к мускарину), открывающие под его действием калиевые каналы, что приводит к гиперполяризации постсинаптической мембраны и генерации ТПСП.
В адренергических синапсах выделяющийся норадреналин может действовать на два типа рецепторов – альфа-адренорецепторы и бета-адренорецепторы. В эффекторной ткани зачастую присутствуют как альфа-, так и бета-адренорецепторы, которые обеспечивают антагонистичекие или синергические действия.
Таким образом, один и тот же медиатор может взаимодействовать с различными рецепторами постсинаптической мембраны и вызывать различные эффекты.
Выделившиеся в синаптическую щель, но не вступившие в реакцию с постсинаптическими рецепторами молекулы медиатора разрушаются с помощью специальных ферментов или обратно всасываются (обратный захват) в пресинаптическое окончание.
Нейронные цепи
Нервная система человека может быть представлена в виде нервной сети, т.е. системы нейронных цепочек, передающих возбуждающие и тормозные сигналы. Нейронные сети построены из трёх главных компонентов: входных волокон, вставочных нейронов и эфферентных нейронов. Самыми простыми и элементарными нейронными цепями являются локальные сети, или микросети. Нередко определённый тип микросети повторяется по всему слою нервной структуры, например коры больших полушарий, и выступает в качестве модуля для особого способа обработки информации.
Локальные сети имеются в разных отделах мозга. Они служат: 1) для усиления слабых сигналов; 2) уменьшения и фильтрации слишком интенсивной активности; 3) выделения контрастов; 4) поддержания ритмов или сохранения рабочего состояния нейронов путём регулировки их входов. Микросети могут оказывать на нейроны-мишени возбуждающее или тормозящее действие.
Локальные сети можно сравнить с интегральными микросхемами в электронике, т.е. стандартными элементами, которые выполняют наиболее часто повторяющиеся операции и могут быть включены в схемы самых разнообразных электронных приборов.
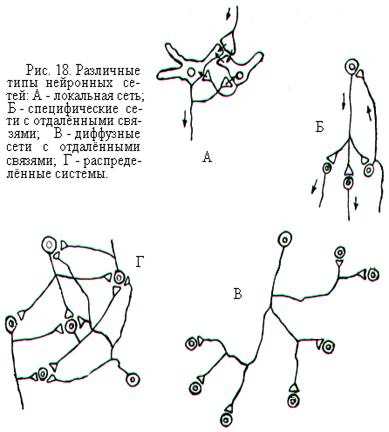 Одну из разновидностей локальных сетей составляют, как правило, нейроны с короткими аксонами (рис. 18). Поэтому задачи и сферы влияния таких нейронов весьма ограничены. Вторую разновидность локальной сети образуют нейроны, достаточно удалённые друг от друга, но принадлежащие к одной нервной области. Основными функциями этих сетей является распространение активности за пределы отдельного модуля или же обеспечение антагонистических взаимодействий между соседними модулями в пределах данной нервной области.
Одну из разновидностей локальных сетей составляют, как правило, нейроны с короткими аксонами (рис. 18). Поэтому задачи и сферы влияния таких нейронов весьма ограничены. Вторую разновидность локальной сети образуют нейроны, достаточно удалённые друг от друга, но принадлежащие к одной нервной области. Основными функциями этих сетей является распространение активности за пределы отдельного модуля или же обеспечение антагонистических взаимодействий между соседними модулями в пределах данной нервной области.
Более сложными являются сети с отдалёнными связями, соединяющие две или несколько областей нервной системы с локальными сетями. Сети с отдалёнными связями могут быть как специфическими, так и диффузными (рис. 18). Специфическое последовательное соединение нескольких областей выполняет функцию передачи информации с периферии в центральную нервную систему (например, проводниковые отделы анализаторов) или с центральных отделов на периферию (например, моторная система). В таких случаях обычно сети с отдалёнными связями называют восходящими и нисходящими путями, или системами. Нервные структуры, входящие в восходящие пути, объединяются по принципу
восходящей иерархии, а образующие нисходящие пути – по принципу нисходящей иерархии.
Самый высокий уровень организации – это система соединений между целым рядом областей, управляющих каким-то поведением, в котором участвует весь организм. Такие сети носят название распределённых систем (рис. 18). Они могут находиться в разных отделах мозга и могут быть связаны гормональными воздействиями или длинными нервными путями. Распределённые системы участвуют в осуществлении высших функций двигательных и сенсорных систем, а также многих центральных систем, обеспечивающих сложные поведенческие акты, абстрактное мышление, речь и другие психофизиологические процессы.
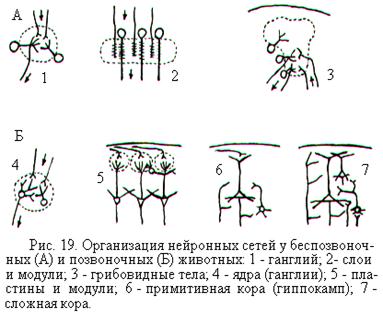 В процессе эволюции происходило усложнение нейронных сетей. У беспозвоночных животных со слабо интегрированной нервной системой нейронные сети обычно организованы либо в форме ганглиев, либо в форме пластинок (рис. 19, А). Ганглии являются структурой с концентрированным расположением синаптических контактов между входными и выходными элементами, а пластинки – структурой с двухслойной организацией таких контактов.
В процессе эволюции происходило усложнение нейронных сетей. У беспозвоночных животных со слабо интегрированной нервной системой нейронные сети обычно организованы либо в форме ганглиев, либо в форме пластинок (рис. 19, А). Ганглии являются структурой с концентрированным расположением синаптических контактов между входными и выходными элементами, а пластинки – структурой с двухслойной организацией таких контактов.
У высших беспозвоночных животных интеграция сигналов на более высоком уровне происходит в нервных центрах, примером чему являются грибовидные тела мозга у насекомых. Грибовидные тела спрятаны в глубине мозга, а не на его поверхности, где они могли бы разрастаться в ширину.
У позвоночных животных и у человека часть нейронных сетей группируются в ганглии. Центры, расположенные в глубине мозга, увеличиваются за счёт образования изгибов, подобно грибовидным телам насекомых. Однако принципиально новая особенность высших позвоночных и человека – это группировка огромного количества нейронов в слои, лежащие на поверхности мозга, т.е. образование коры (рис. 19, Б).
Кора расположена таким образом, что нейроны всех её слоёв доступны для любых входных сигналов. Совместно с локальными сетями, образованными ответвлениями отростков нейронов и вставочными нейронами, кора обладает огромными возможностями для интеграции, хра-
нения и комбинирования информации. В каждом участке, или поле коры, многократно повторяются однотипные модули (локальные сети), благодаря которым данное поле способно осуществлять специфические операции с участием определённых входных и выходных связей (зрительное поле, слуховое поле). При переходе к соседнему полю коры все эти три элемента, т.е. локальные сети, входы и выходы, слегка изменяются. Изменяются и функциональные свойства. Таким образом, каждое из полей коры является участком, приспособленным для выполнения определённых функций в той распределённой системе, в состав которой оно входит.
ОСНОВНЫЕ ЗАКОНЫ ФУНКЦИОНИРОВАНИЯ
НЕРВНЫХ СЕТЕЙ
Дивергенция и конвергенция путей
Во всех изученных нервных сетях обнаружены дивергенция и конвергенция путей. Дивергенцией называют способность нейрона устанавливать многочисленные синаптические связи со многими другими нервными клетками (рис. 20, А). Так, например аксон чувствительного нейрона входит в задние рога спинного мозга в составе задних корешков и в спинном мозге разветвляется на множество веточек (коллатералей), образующих синапсы на многих вставочных нейронах и мотонейронах. Благодаря процессу дивергенции одна и та же нервная клетка может участвовать в различных нервных реакциях и контролировать большое число других нейронов. Такое расширение сферы действия и распространение сигнала в нервных сетях называется иррадиацией. Иррадиировать может как возбуждение так и торможение.
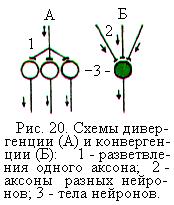 Схождение многих нервных путей к одному и тому же нейрону носит название конвергенции (рис. 20, Б). Например, на каждом мотонейроне спинного мозга образуют синапсы тысячи отростков чувствительных, а также возбуждающих и тормозных вставочных нейронов из разных отделов ЦНС. Вследствие конвергенции многих нервных путей к одному нейрону этот нейрон осуществляет интеграцию одновременно поступающих по разным путям возбуждающих и тормозных сигналов. Если в результате алгебраического сложения ВПСП и ТПСП, возникающих на мембране нейрона, возбуждение будет преобладать, то нейрон возбудится и пошлёт нервный импульс ко второй клетке. Если будет преобладать достаточной величины ТПСП, то нейрон затормозится. Такое сложение постсинаптических потенциалов называют пространственной, или одновременной суммацией.
Схождение многих нервных путей к одному и тому же нейрону носит название конвергенции (рис. 20, Б). Например, на каждом мотонейроне спинного мозга образуют синапсы тысячи отростков чувствительных, а также возбуждающих и тормозных вставочных нейронов из разных отделов ЦНС. Вследствие конвергенции многих нервных путей к одному нейрону этот нейрон осуществляет интеграцию одновременно поступающих по разным путям возбуждающих и тормозных сигналов. Если в результате алгебраического сложения ВПСП и ТПСП, возникающих на мембране нейрона, возбуждение будет преобладать, то нейрон возбудится и пошлёт нервный импульс ко второй клетке. Если будет преобладать достаточной величины ТПСП, то нейрон затормозится. Такое сложение постсинаптических потенциалов называют пространственной, или одновременной суммацией.
Афферентных нейронов в нервной системе примерно в 5 раз больше, чем эфферентных. В связи с этим многие афферентные импульсы поступают к одним и тем же вставочным и эфферентным нейронам, которые являются для импульсов общими конечными путями к рабочим органам.
Впервые закономерности общих конечных путей были изучены в начале 20-го века английским физиологом Ч. Шеррингтоном. Благодаря общим конечным путям одна и та же ответная рефлекторная реакция определённой группы мотонейронов может быть получена при раздражении различных нервных структур. Рефлексы, дуги которых имеют общий конечный путь, подразделяются на союзные и антагонистические. Встречаясь на общих конечных путях, союзные рефлексы взаимно усиливают друг друга, а антагонистические – тормозят друг друга, как бы конкурируя за захват общего конечного пути. Преобладание на конечных путях той или иной, в том числе и поведенческой, рефлекторной реакции обусловлено её значением для жизнедеятельности организма в данный момент.
Временнáя и пространственная суммация. Окклюзия
Если ритмическое раздражение наносится с достаточно большой частотой, то импульсы, поступающие друг за другом в синапс по одному нервному пути, вызывают в постсинаптическом нейроне быстро возникающие друг за другом ВПСП. Эти ВПСП суммируются, достигая в конечном итоге порогового уровня, что приводит к развитию потенциала действия, или нервного импульса, в постсинаптическом нейроне. Такого рода повышение возбудимости нейрона в ходе последовательных ВПСП называется временнóй суммацией, или временным облегчением. Временнáя суммация играет очень важную физиологическую роль, потому что многие нейронные процессы (например, разряд рецептора) имеют ритмический характер и, таким образом, могут суммироваться, давая начало надпороговому возбуждению в постсинаптической мембране.
 Наличие конвергенции многих путей, т.е. нервных цепочек, на одних и тех же эфферентных нейронах лежит в основе пространственной суммации и окклюзии. Пространственная суммация, или пространственное облегчение, наблюдается в том случае, когда одновременно по нескольким нервным путям поступают слабые импульсы к одному и тому же нейрону. Раздельная стимуляция каждого из этих входов к нейрону вызывает только подпороговый ВПСП. При одновременной же стимуляции нескольких входов ВПСП суммируются и в нейроне возникает потенциал действия.
Наличие конвергенции многих путей, т.е. нервных цепочек, на одних и тех же эфферентных нейронах лежит в основе пространственной суммации и окклюзии. Пространственная суммация, или пространственное облегчение, наблюдается в том случае, когда одновременно по нескольким нервным путям поступают слабые импульсы к одному и тому же нейрону. Раздельная стимуляция каждого из этих входов к нейрону вызывает только подпороговый ВПСП. При одновременной же стимуляции нескольких входов ВПСП суммируются и в нейроне возникает потенциал действия.
Окклюзия довольно часто обнаруживается при возбуждении группы мотонейронов. При окклюзии ответ на одновременную стимуляцию нескольких нервных путей к мотонейронам меньше, чем сумма ответов, наблюдающихся при раздельной стимуляции этих путей. Причина окклюзии состоит в том, что часть афферентных путей (на рис.21 ответвления аксонов 1 и 2) вследствие конвергенции подходит к одним и тем же мотонейронам (б). При раздельной стимуляции входов 1 и 2 мотонейрон б будет возбуждаться дважды: сначала совместно с нейроном а и затем совместно с нейроном в. При одновременной стимуляции входов 1 и 2 нейрон б будет возбуждаться только один раз. В этом случае рефлекторный ответ будет меньше алгебраической суммы ответов при раздельной стимуляции.
Таким образом, окклюзия – это явление, противоположное пространственной суммации. При увеличении интенсивности стимуляции входов к группе нейронов пространственная суммация может перейти в окклюзию.
Торможение в нервных сетях
Торможением называют нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения, в результате чего уменьшается или совсем отсутствует ответная реакция на раздражение. Явление торможения в нервных центрах было впервые открыто И.М. Сеченовым в опытах на лягушке и описано им в книге «Рефлексы головного мозга». И.М. Сеченов опускал лапку лягушки в кислоту и одновременно раздражал зрительные бугры промежуточного мозга, накладывая на них кристаллик поваренной соли. Он наблюдал резкую задержку и даже полное отсутствие спинномозгового рефлекса отдёргивания лапки из кислоты. После снятия кристаллика соли и промывания мозга физиологическим раствором время рефлекса, т.е. скорость вынимания лапки из кислоты, постепенно восстанавливалось. Этот опыт вошёл в историю физиологии под названием Сеченовское торможение. Он доказывает, что торможение – это активный процесс, осуществляющийся в нервной системе наряду с торможением.
Тормозные процессы – необходимый компонент в координации нервной деятельности. Во-первых, процесс торможения ограничивает распространение возбуждения на соседние нервные центры; во-вторых, процесс торможения выключает деятельность не нужных в данный момент центров и органов; в-третьих, развитие торможения в нервных центрах предохраняет их от чрезмерного перенапряжения при работе, т.е. играет охранительную роль.
В нервных сетях обнаружено несколько видов торможения. Их классифицируют по характеру, механизму и месту возникновения. По характеру возникновения различают первичное и вторичное торможение. Первичное торможение нейрона возникает под влиянием раздражения сразу без предварительного возбуждения этого нейрона и осуществляется с участием тормозных синапсов. Вторичное торможение осуществляется без участия специальных тормозных структур и возникает вследствие перехода возбуждения в торможение. По механизму возникновения торможение может быть гиперполяризационным и деполяризационным, а по месту возникновения – постсинаптическим и пресинаптическим.
Постсинаптическое торможение
Постсинаптическое торможение имеет широкое распространение в ЦНС. Оно возникает в постсинаптической мембране нейрона в результате действия тормозного медиатора и связано с наличием в нейронных цепях специальных тормозных нейронов. Различают прямое и возвратное постсинаптическое торможение.
Прямое постсинаптическое торможение характерно для мотонейронов. При этом виде торможения нервные импульсы из высших отделов ЦНС поступают к вставочным тормозным нейронам и вызывают у них деполяризацию мембраны и обычный процесс возбуждения. Возбудившись, вставочный тормозной нейрон посылает по аксону нервный импульс к мотонейрону. Особенностью тормозного нейрона является то, что окончания его аксона выделяют не возбуждающий, а тормозной медиатор, например, гамма-аминомасляную кислоту, глицин и др. Под влиянием тормозного медиатора происходит кратковременная гиперполяризация постсинаптической мембраны мотонейрона и возникает ТПСП. В результате мотонейрон тормозится, импульсы от него не поступают к мышечным волокнам и они не сокращаются.
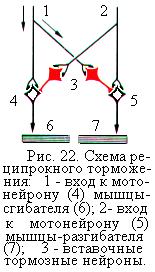 Таким образом, вставочные тормозные нейроны не возбуждают, а тормозят те клетки, на которых оканчиваются их аксоны. Вставочные тормозные нейроны обусловливают осуществление антагонистического, или реципрокного, торможения (рис. 22). Реципрокным торможением является торможение мотонейронов мышц-антагонистов, например, мышц-сгибателей и мышц-разгибателей. При активации нервных путей, возбуждающих мотонейроны мышц-сгибателей, мотонейроны мышц-разгибателей тормозятся импульсами тормозных вставочных нейронов, и наоборот.
Таким образом, вставочные тормозные нейроны не возбуждают, а тормозят те клетки, на которых оканчиваются их аксоны. Вставочные тормозные нейроны обусловливают осуществление антагонистического, или реципрокного, торможения (рис. 22). Реципрокным торможением является торможение мотонейронов мышц-антагонистов, например, мышц-сгибателей и мышц-разгибателей. При активации нервных путей, возбуждающих мотонейроны мышц-сгибателей, мотонейроны мышц-разгибателей тормозятся импульсами тормозных вставочных нейронов, и наоборот.
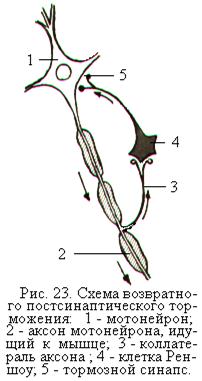
Возвратное постсинаптическое торможение – это торможение нейронов собственными импульсами, поступающими по возвратным коллатералям (боковым ответвлениям аксона) к специальным тормозным клеткам, аксоны которых образуют тормозные синапсы на этих же нейронах. Такими специальными тормозными нейронами являются клетки Реншоу в спинном мозге, клетки Пуркинье мозжечка, корзинчатые клетки в промежуточном мозге и мозжечке.
Клетки Реншоу участвуют в регуляции уровня активности отдельных мотонейронов. При возбуждении мотонейрона спинного мозга импульсы поступают по его аксону к мышечным волокнам и одновременно по коллатерали аксона – к тормозной клетке Реншоу. Аксон клетки Реншоу «возвращается» к этому же мотонейрону и, выделяя тормозной медиатор, вызывает его торможение. Чем больше возбуждающих импульсов
посылает мотонейрон на периферию, в том числе и к тормозной клетке Реншоу, тем сильнее возвратное торможение (рис. 23). Такая замкнутая система действует как механизм саморегуляции мотонейрона, ограничивая степень его возбуждения и предохраняя от чрезмерного ритма, что важно для нормальной работы двигательного аппарата. Эту же роль играет возвратное торможение и в других нервных сетях.
 Клетки Пуркинье мозжечка своими тормозящими влияниями на клетки подкорковых ядер и ствола мозга участвуют в регуляции тонуса мышц.
Клетки Пуркинье мозжечка своими тормозящими влияниями на клетки подкорковых ядер и ствола мозга участвуют в регуляции тонуса мышц.
Корзинчатые клетки промежуточного мозга и мозжечка вызывают синхронное торможение большого количества связанных с ними нейронов (рис. 24), регулируя поступление импульсов через эти нейроны в другие отделы ЦНС, в том числе и в кору больших полушарий.
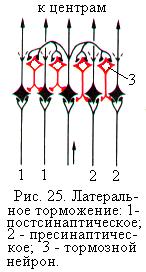 Ещё одной формой возвратного торможения является латеральное торможение (рис. 25). Латеральное торможение осуществляется тормозными вставочными нейронами, которые соединены таким образом, что они влияют не только на возбуждённую клетку, но и на соседние клетки с такими же функциями, в которых возбуждение отсутствует или является более слабым. В результате в этих соседних клетках развивается очень глубокое торможение.
Ещё одной формой возвратного торможения является латеральное торможение (рис. 25). Латеральное торможение осуществляется тормозными вставочными нейронами, которые соединены таким образом, что они влияют не только на возбуждённую клетку, но и на соседние клетки с такими же функциями, в которых возбуждение отсутствует или является более слабым. В результате в этих соседних клетках развивается очень глубокое торможение.  Торможение такого типа называется латеральным, потому что образующаяся зона торможения находится сбоку, т.е. латерально, по отношению к возбуждённому нейрону. Возбуждённый нейрон оказывается со всех сторон окружённым зоной торможения. Латеральное торможение обеспечивает контраст, т.е. выделение существенных сигналов из фона. Оно играет особенно важную роль в сенсорных системах, где может иметь форму постсинаптического торможения или пресинаптического.
Торможение такого типа называется латеральным, потому что образующаяся зона торможения находится сбоку, т.е. латерально, по отношению к возбуждённому нейрону. Возбуждённый нейрон оказывается со всех сторон окружённым зоной торможения. Латеральное торможение обеспечивает контраст, т.е. выделение существенных сигналов из фона. Оно играет особенно важную роль в сенсорных системах, где может иметь форму постсинаптического торможения или пресинаптического.
Пресинаптическое и пессимальное торможение
Торможение может развиваться ещё в пресинаптическом отделе путём угнетения процесса высвобождения медиатора возбуждающими нервными окончаниями. В этом случае свойства постсинаптической мем-
браны не изменяются. Пресинаптическое торможение обнаружено в различных отделах ЦНС, но наиболее часто оно встречается в мозговом стволе и особенно в спинном мозге.
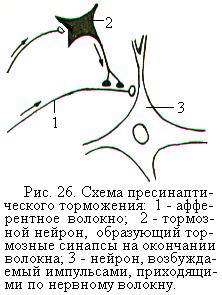 Пресинаптическое торможение возникает в пресинаптической области перед синаптическим контактом, т.е. там, где аксон возбуждающего нейрона лишён миелиновой оболочки перед переходом его в нервное окончание (рис. 26). С этим лишённым миелина участком контактирует аксон тормозного вставочного нейрона и образует тормозной аксо-аксональный синапс. Импульсы, поступающие по аксону тормозного нейрона в аксо-аксональный синапс, вызывают выделение тормозного медиатора (вероятнее всего гамма-аминомасляной кислоты), который вызывает сильную деполяризацию окончания аксона возбуждающего нейрона. Деполяризованный участок аксона теряет способность проводить нервные импульсы к синапсу и, следовательно, ко второй нервной клетке.
Пресинаптическое торможение возникает в пресинаптической области перед синаптическим контактом, т.е. там, где аксон возбуждающего нейрона лишён миелиновой оболочки перед переходом его в нервное окончание (рис. 26). С этим лишённым миелина участком контактирует аксон тормозного вставочного нейрона и образует тормозной аксо-аксональный синапс. Импульсы, поступающие по аксону тормозного нейрона в аксо-аксональный синапс, вызывают выделение тормозного медиатора (вероятнее всего гамма-аминомасляной кислоты), который вызывает сильную деполяризацию окончания аксона возбуждающего нейрона. Деполяризованный участок аксона теряет способность проводить нервные импульсы к синапсу и, следовательно, ко второй нервной клетке.
Пресинаптическое торможение особенно эффективно при обработке информации, поступающей к нейрону по различным пресинаптическим путям. В этом случае возбуждение, поступающее по одному из синаптических входов, может быть избирательно уменьшено или даже полностью прекращено при отсутствии влияния на другие входы к нейрону. Сам постсинаптический нейрон остаётся незаторможенным и способен реагировать на импульсы, поступающие по другим входам. Пресинаптическое торможение обусловливает ограничение притока афферентных импульсов к нервным центрам.
Торможение деятельности нейрона может осуществляться и без участия особых тормозных структур. В этом случае торможение развивается в возбуждающих синапсах в результате сильной деполяризации мембраны под влиянием слишком частого поступления нервных импульсов в пресинаптический отдел. Такой тип торможения был изучен русским физиологом Н.Е. Введенским и назван пессимальным.
В основе развития пессимального торможения лежит стойкая деполяризация, которая может возникнуть как в постсинаптической, так и в пресинаптической мембране синапса под влиянием очень частого поступления сильных импульсов. К пессимальному торможению особенно склонны вставочные нейроны спинного мозга, нейроны ретикулярной формации и некоторые другие нервные клетки, в которых при частом ритмическом раздражении деполяризация синаптических мембран может быть сильной и стойкой.
СПИННОЙ МОЗГ
Строение спинного мозга
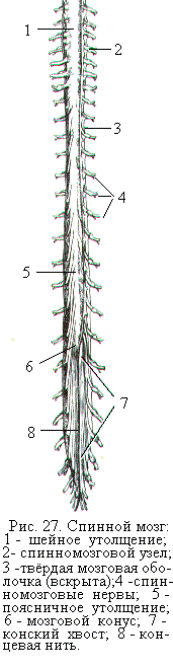 Спинной мозг – наиболее древний в филогенетическом плане отдел ЦНС. Он располагается в позвоночном канале и представляет собой немного сплюснутый в переднезаднем направлении цилиндрический тяж длиной около 45 см у взрослого человека (рис. 27). Спинной мозг покрыт тремя оболочками – твёрдой, паутинной и мягкой. Внутри позвоночного канала он удерживается при помощи связок, которые идут от твёрдой мозговой оболочки к внутренней стенке канала. Пространство между твёрдой и паутинной (субарахноидальное пространство), а также между паутинной и мягкой оболочками (субдуральное пространство) заполнено спинномозговой жидкостью.
Спинной мозг – наиболее древний в филогенетическом плане отдел ЦНС. Он располагается в позвоночном канале и представляет собой немного сплюснутый в переднезаднем направлении цилиндрический тяж длиной около 45 см у взрослого человека (рис. 27). Спинной мозг покрыт тремя оболочками – твёрдой, паутинной и мягкой. Внутри позвоночного канала он удерживается при помощи связок, которые идут от твёрдой мозговой оболочки к внутренней стенке канала. Пространство между твёрдой и паутинной (субарахноидальное пространство), а также между паутинной и мягкой оболочками (субдуральное пространство) заполнено спинномозговой жидкостью.
Вверху над первым шейным позвонком – атлантом – спинной мозг переходит в продолговатый мозг, а внизу на уровне 2-го поясничного позвонка заканчивается заострением – мозговым конусом. От конуса отходит концевая нить, прикрепляющая снизу спинной мозг ко 2-му копчиковому позвонку. Спинной мозг имеет два утолщения – шейное и поясничное, образованные большим скоплением нейронов и нервных волокон. Из этих утолщений выходят нервы к верхним и нижним конечностям.
|
|
Дата добавления: 2014-12-26; Просмотров: 1165; Нарушение авторских прав?; Мы поможем в написании вашей работы!