КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Основы межклеточного взаимодействия в организме. Физиология синапсов
|
|
|
|
Классификация и свойства нервных волокон. Законы проведения возбуждения по нервным волокнам
Возбудимые клетки как проводники электричества. Распространение электротонического потенциала. Константа длины. Механизм проведения возбуждения по немиелинизированным и миелинизированным нервным волокнам
При генерации потенциала действия
А – потенциал действия, Б – изменение возбудимости
Лабильность, или функциональная подвижность, измеряется в количестве потенциалов действия, которое может воспроизвести ткань за единицу времени при ритмическом раздражении. Она зависит от длительности ПД, а, следовательно, от периода абсолютной рефрактерности. Для нервной ткани она в среднем равна 1000 импульсам в секунду, для мышечной – 200-300 имп/c, а для нервно-мышечного синапса – 100-150 имп/c.
Мембрана нейрона и его отростков (нервных волокон) обладает способностью не только генерировать потенциал действия, но и передавать го на относительно большие расстояния без уменьшения его амплитуды и расходования энергии, поэтому нервные импульсы распространяются без затухания (бездекрементно). Способность к распространению возбуждения связана с тем, что во время потенциала действия происходит изменение знака заряда в возбужденном участке мембраны (инверсия заряда). Между ним и невозбужденными соседними участками мембраны возникают локальные электрические токи, под действием которых происходит деполяризация новых соседних участков, что приводит к формированию в них потенциала действия. За счет последовательного охвата возбуждением все новых участков нервного волокна возбуждение перемещается вдоль дендритов и аксонов. Чем больше диаметр волокна, тем больше скорость распространения возбуждения.
|
|
|
Особенности проведения возбуждения в нервных волокнах определяются наличием или отсутствием у них оболочки. Нервные волокна делятся на миелиновые и безмиелиновые, то есть покрытые особой миелиновой оболочкой или не покрытые. Эта оболочка образуется на периферии шванновскими клетками, а в головном мозге – клетками олигодендроглии.
Механизм проведения импульса по нервным волокнам объясняется теорией “местных токов” Тасаки (рисунок 3 и рисунок 4). В безмиелиновом нервном волокне ПД генерируется каждой точкой поверхности мембраны – так называемое непрерывное проведение возбуждения. Во время пикового ПД на наружной поверхности волокна происходит перезарядка, в результате чего возникает местный ток от соседнего положительно заряженного участка к возбужденному, а внутри – наоборот. Таким образом образуется кольцевой ток.
В миелиновых волокнах слой миелина не целиком охватывает волокно, а имеет промежутки – перехваты Ранвье. Сам миелин обладает высоким сопротивлением электрическому току, поэтому в участках, покрытых миелином, потенциал действия генерироваться не может. Перехваты Ранвье располагаются примерно через каждые 0,5-1,5 мм (в зависимости от толщины оболочки), в них обнаружено очень много (около 12 тыс/мкм2) натриевых каналов, поэтому в миелиновых волокнах проведение возбуждения происходит скачкообразно (сальтаторно).
Между соседними участками возникает большой по величине ток (ПД = 100-120 мВ), который в 3-5 раз превышает порог раздражения и способен возбудить соседние участки, что составляет фактор надежности.
Скорость проведения возбуждения пропорциональна корню квадратному от диаметра волокна. Самые тонкие волокна – безмиелиновые, а в миелиновых волокнах чем толще слой миелина, тем больше расстояние между перехватами Ранвье. В безмиелиновых волокнах местные кольцевые токи последовательно распространяются “маленькими шажочками” непрерывно, скорость проведения возбуждения в безмиелиновых волокнах гораздо ниже, чем в миелиновых. Такая передача импульса является медленной и неэкономичной в плане энергозатрат. Поэтому безмиелиновыми являются только самые тонкие волокна (менее 1 мкм в диаметре), проводящие возбуждение к медленно работающим органам – кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре.
|
|
|
Константа длины – это расстояние, на котором местный ток сохраняет пороговое значение, то есть способен вызывать образование ПД на следующем участке нервного волокна.
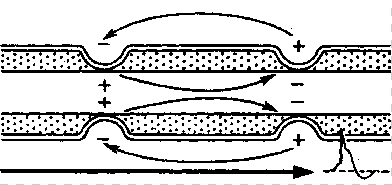
Рисунок 3 – Передача возбуждения в миелинизированном нервном волокне
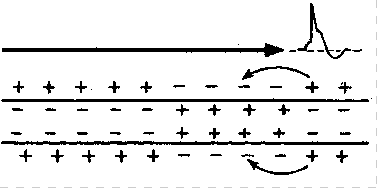
Рисунок 4 – Передача возбуждения в немиелинизированном нервном волокне
Такая передача импульса является медленной и неэкономичной в плане.
Все нервные волокна делят на группы А, В и С. Группы А и В образованы миелиновыми волокнами, С – немиелинизированными.
Классификация нервных волокон по Эрлангеру и Гассеру, Ллойду представлена в таблице 2.
Таблица 2
Классификация нервных волокон
| Тип волокна | скорость проведения, м/с | Диаметр, мкм | Наличие миелиновой оболочки | Локализация, функции |
| Аa | 70-120 м/c | 12-22 | есть | проводят возбуждение от тактильных рецепторов кожи, а также от спинальных α-мотонейронов к сократительным волокнам скелетных мышц |
| Аb | 40-70 м/с | 8-12 | есть | от спинальных α-мотонейронов к сократительным волокнам скелетных мышц |
| Аg | 15–30 м/с | 4-8 | есть | от спинальных γ-мотонейронов к сократительным клеткам мышечных веретен |
| Аd | 5-15 м/с | 1-4 | есть | от спинальных γ-мотонейронов к сократительным клеткам мышечных веретен |
| В | 3-15 м/с | 1-3 | есть | в вегетативных преганглионарных волокнах |
| С | 0,5-3 | 0,5-2 | нет | некоторые интерорецепторы, болевые и температурные рецепторы кожи, постганглионарные вегетативные волокна |
Свойства нервных волокон:
1. Самая высокая возбудимость по сравнению с мышечной и железистой тканями.
2. Наибольшая лабильность по сравнению с мышечной и железистой тканями.
3. Нервные волокна практически не утомляются, так как основной процесс возбуждения идет без расхода энергии за счет пассивных входа Na+ и выхода К+ в малых количествах на фоне очень больших градиентов их концентраций.
|
|
|
4. Большая скорость прохождения импульса.
5. Наличие от нейрона к периферии (к окончанию аксона) аксонного тока, с которым в нервные окончания поступают все необходимые вещества.
Законы проведения возбуждения по нервным волокнам:
1) физиологическая целостность – проведение нервного импульса возможно только при полной анатомической целостности волокна;
2) изолированное проведение возбуждения – потенциал действия не передается с одного нервного волокна на другое;
3) двустороннее проведение возбуждения – возбуждение может идти как центробежно, так и центростремительно.
Переход возбуждения от нейрона к нейрону или к мышечной или железистой клетке происходит в местах особых контактов – синапсов (Термин «синапс» ввёл Ч. Шеррингтон в 1897 г.). Синапс – это место контакта двух возбудимых клеток (в частности, нервного окончания и мышцы, нервного окончания одного нейрона и тела другого (вставочного) нейрона, нервного окончания и железистой клетки), обеспечивающее передачу нервных импульсов с одной возбудимой клетки на другую. В своем составе синапс имеет пре- и постсинаптическую мембраны, между которыми находится синаптическая щель. Синапсы могут быть аксодендритные (между окончанием аксона и дендритом следующего нейрона), аксосоматические, реже – сома-соматические, дендродендритные и дендросоматические.

Классификация синапсов:
1. По виду соединяемых клеток:
- межнейронные синапсы – находятся в ЦНС и вегетативных ганглиях;
- нервно-мышечные синапсы - соединяют аксоны мотонейрона с мышечным волокном.
2. По эффекту:
- возбуждающие, т.е. запускающие генерацию ПД;
- тормозные, т.е. препятствующие возникновению ПД.
3. По способу передачи сигнала:
- химические синапсы – передача осуществляется с помощью химического
посредника – медиатора;
- электрические синапсы – ПД непосредственно (электротонически) переда-
|
|
|
ется на постсинаптическую клетку;
- смешанные синапсы – наряду с химической передачей имеются участки с
электротоническим механизмом передачи (например, в реснитчатом
ганглии птиц, спинном мозге лягушки).
4. По природе нейромедиатора:
- холинергические (медиатор – ацетилхолин);
- адренергические (норадреналин);
- дофаминергические (дофамин);
- ГАМКергические (ГАМК);
- глутаматергические (глутамат);
- серотонинергические (серотонин);
- пептидергические (пептиды);
- глицинергические (глицин).
6. По местоположению:
- центральные (головной и спинной мозг);
- периферические.
В электрических синапсах возбуждение передается электротонически, за счет локальных круговых токов между пре- и постсинаптической мембранами. Ширина синаптической щели в них составляет 1-2 нм, между синаптическими мембранами существуют узкие щелевые контакты, обладающие низким электрическим сопротивлением. В них почти нет утечки через внеклеточную среду, поэтому изменения потенциала в пресинаптической мембране могут эффективно передаваться на постсинаптическую мембрану, в которой под действием потенциалов действия с пресинаптической мембраны меняется ионная проницаемость и генерируются свои потенциалы действия.
Электрические синапсы встречаются чаще у беспозвоночных и низших позвоночных (например, у ракообразных). В стволе мозга млекопитающих электрические синапсы имеются в ядрах тройничного нерва и в нижней оливе мозгового ствола.
В электрических синапсах проведение возбуждения происходит очень быстро, практически без синаптической задержки, ток возможен в обоих направлениях (но легче – в одном, от пресинаптического окончания к постсинаптической мембране). Электрические синапсы дают возможность получать постоянные, повторяющиеся реакции и синхронизировать активность многих нейронов.
|
|
|
|
|
Дата добавления: 2014-12-23; Просмотров: 1358; Нарушение авторских прав?; Мы поможем в написании вашей работы!