КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Пищеварительная и нервная система. Типы ротовых аппаратов. Пищеварительная система
|
|
|
|
Line, семейство L1
Ретроэлементы без длинных концевых повторов
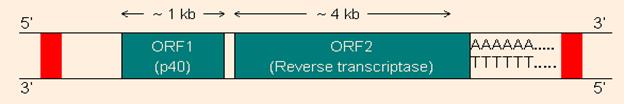
ORF 1 кодирует РНК-связывающий белок, ORF 2 обратную транскриптазу и эндонуклеазу. Красные- прямые повторы, которые есть у всех ретроэлементов
ЕЩЕ РАЗ:
SINEs
Short Interspersed Nuclear Elements
Не содержат функциональных генов (LINE элементы
с делетированной внутренней частью).
У человека –Alu- элементы.
Три отдела: передняя, средняя и задняя кишка. Между передней и средней кишкой находится кардиальный клапан, средней и задней кишкой–пилорический. Передняя кишка представлена глоткой, пищеводом, зобом, механическим желудком. В зависимости от потребляемой пищи возможны вариации в строении: отсутствует зоб, желудок. Зоб – место временного пребывания пищи, отчасти здесь происходит пищеварение; функция желудка –размельчение (перетирание) пищи. Глотка у насекомых, питающихся жидкой пищей, мускулистая, выполняет функцию насоса. Слюнные железы открываются в ротовую полость, как правило, около основания нижней губы. Содержащиеся в слюне ферменты обеспечивают начальные этапы пищеварения. У кровососущих насекомых слюна содержит вещества, препятствующие
свертыванию крови – антикоагулянты. В некоторых случаях слюнные железы меняют свою функцию (у гусениц превращаются в прядильные железы). В средней (тонкой) кишке происходит переваривание и всасывание пищи. В начальный участок кишки у некоторых насекомых (таракан и др.) впадает
несколько слепых выпячиваний кишечника – пилорические придатки – они увеличивают всасывающую поверхность. Стенки средней кишки образуют складки – крипты. Тип пищеварительных ферментов зависит от пищевого режима насекомых. Секреция ферментов у насекомых голокринная и мерокринная. Эпителий средней кишки у многих насекомых выделяет вокруг содержимого кишки перитрофическую мембрану, роль которой важна в процессах переваривания и всасывания пищевых веществ. Кроме того, она пре-
|
|
|
дохраняет эпителий средней кишки от механических повреждений. Задняя (прямая) кишка нередко отличается значительной длиной и подразделяется на несколько участков. Здесь у большинства насекомых находятся ректальные железы. Функции отдела: формирование и удаление экскрементов, всасывание воды из пищевой массы, переваривание пищи с помощью симбионтов (характерно для личинок некоторых видов насекомых). Отделы кишечника разделены клапанами, препятствующими обратному току пищи. Перед-
ний и средний отделы разделяет кардиальный клапан, средний и задний – пилорический. Тип пищи накладывает отпечаток на морфологию органов питания. Пища насекомых разнообразная и охватывает почти все вещества животного и растительного происхождения. По пищевой специализации насекомые
подразделяются на следующие группы: пантофаги, полифаги, олигофаги и монофаги. Классификация насекомых по пищевым режимам: фитофаги, зоофаги, сапрофаги, копрофаги, некрофаги, гематофаги. Нервная система: Нервная система, как у всех членистоногих, брюшная нервная цепочка. Она подразделяется на центральную, периферическую и симпатическую. Центральная нервная система состоит из головного мозга, нервной цепочки, начинающейся подглоточным ганглием. Головной мозг имеет 3 отдела. В протоцеребруме хорошо развиты зрительные доли, внутри полушарий размещаются грибовидные тела – ассоциативные центры, они хорошо развиты у общественных насекомых. Дейтоцеребрум иннервирует усики, основная часть его дифференцирована в обонятельные доли. Тритоцеребрум иннервирует верхнюю губу, от него берет начало симпатическая система. Подглоточный ганглий иннервирует ротовые конечности. У большинства насекомых ганглии брюшной нервной цепочки концентрируются в продольном направлении. Периферическая нервная система образована нервами, отходящими от центральной и симпатической системы, а также нейронами, расположенными по всему телу. Симпатическая система состоит из трех отделов: стоматогастрического, брюшного и хвостового. Имеются нейросекреторные клетки во всех отделах нервной системы, выделяемые секреты транспортируются в железы внутренней секреции (прилежащие тела, кардиальные тела, проторакальные железы). Органы чувств: Морфологической и функциональной основой органов чувств являются
|
|
|
сенсиллы, разбросанные по телу насекомых поодиночке или собранные в скопления (органы зрения, слуха и т. п.). Каждая сенсилла состоит из нескольких элементов. Органы чувств подразделяют на механо-, хемо-, фоторецепторы и другие. Механорецепторы воспринимают механическое раздражение. Они представлены
либо отдельными сенсиллами (трихоидные, колоколовидные, хордотональные и др.), либо органами (Джонстонов, тимпанальные). Хеморецепторы – волоски, пластинки, ямки. Фоторецепторы – глаза. Для насекомых характерны как сложные глаза, так и простые. Сложные глаза состоят из омматидиев, число которых варьируется у разных представителей. Каждый омматидий имеет оптическую и сенсорную часть, строение которых сходно с устройством глаз ракообразных. Сложные глаза дневных и ночных насекомых в строении отличаются. Тип глаз первых оппозиционный, вторых – суперпозиционный. В первом случае пигмент, экранизирующий омматидии, неподвижен, во втором – пигмент способен перемещаться,
скапливаясь в верхней части глаза. Простые глаза (1–3) характерны для некоторых отрядов насекомых, они иннервируются от средней части протоцеребрума, а не от зрительных долей. У личинок насекомых с полным превращением имеются латеральные глазки (стеммы), иннервация их идет от зрительных долей мозга. Помимо перечисленных органов чувств, у насекомых имеются рецепторы влажности, терморецепторы, органы равновесия. Типы ротовых аппаратов: Исходным типом ротового аппарата является грызущий (ортоптероидный). Он встречается у насекомых многих отрядов (таракановые, прямокрылые, стрекозы, жуки и др.). В его состав входят следующие элементы: верхняя губа, мандибулы, максиллы, нижняя губа и гипофаринкс. Лакающий (пчелы, шмели) образован верхней губой, мандибулами, в максиллах развита и удлинена наружная жевательная лопасть (galea), которая образует верхнюю и часть боковой поверхности хоботка, нижняя губа представлена удлиненным щупиком (palpi), который образует нижнюю и часть боковой поверхности хоботка. Внутри хоботка размещается язычок, образованный внутренними (glossae) лопастями нижней губы. Сосущий ротовой аппарат (чешуекрылые) включает верхнюю губу, у немногих предста вителей (зубатые моли) мандибулы, нижнюю губу в виде небольшой площадки со щупиками, хоботок образован удлиненными наружными жевательными лопастями максилл. Колюще-сосущий ротовой аппарат (комары, клопы) в своем составе имеет весь набор ротовых конечностей, но они утратили исходную форму, большая часть превратилась в стилеты, служащие для прокалывания покровов животных и растений. Нижняя губа в этом аппарате выполняет функцию футляра. Лижущий (фильтрующий) ротовой аппарат характерен для мух, в нем хорошо развиты лабеллумы нижней губы, мандибулы и максиллы отсутствуют.
|
|
|
90. Кровеносная и дыхательная системы насекомых. Дыхательная система: Органами дыхания являются трахеи. По бокам тела находится до 10 пар дыхалец. По числу и месту расположения дыхалец насекомых делят на 3 группы: голопнейстические, гемипнейстические и апнейстические. Стигмы ведут в атриальную полость, стенки которой образуют замыкательный аппарат и систему фильтрации воздуха, и далее в систему трахей. Трахеи многократно ветвятся и опутывают все органы. Заканчивается каждая
трахея концевой клеткой с радиально расходящимися отростками. Концевые веточки этой клетки (трахеолы) проникают внутрь отдельных клеток тела насекомого. У многих насекомых имеются воздушные мешки. В отличие от трахей в них нет тенидий. У малоактивных насекомых, обитающих в условиях повышенной влажности, поступление кислорода в трахейную систему и удаление из нее углекислого газа происходит путем диффузии через постоянно открытые дыхальца. У большинства насекомых дыхание активное, происходит расслабление и сжатие брюшка. Дыхательные системы, имеющие стигмы, называются открытыми. Насекомые имеют и закрытые системы, для которых характерно отсутствие дыхалец, воздух в организм поступает в этом случае через трахейные, ректальные, дыхальцевые жабры или через покровы. Форма трахейных жабр пластинчатая, кустистая, они представляют собой тонкостенные выросты, пронизанные сетью трахей. Место положения трахейных жабр варьируется. Функции трахей: проводники воздуха, внутренний скелет, поддерживающий внутренние органы в определенном положении.
|
|
|
Кровеносная система: Незамкнутая. Полость тела с открытой кровеносной системой называется гемоцель. В связи с особенностями строения дыхательной системы эта система у насекомых развита сравнительно слабо. Сердце размещено в брюшке, оно поделено на камеры (число их варьируется у разных насекомых), имеются остии с клапанами. От сердца отходит аорта, которая открывается свободно. Совместная работа сердца и диафрагм обеспечивает циркуляцию гемолимфы по телу насекомого. Движение гемолимфы в крыльях осуществляется по жилкам. Имеются местные пульсирующие органы в виде ампулы, мембра-
нозной сократительной перепонки со специальной мышцей. Подобные образования располагаются в основании усиков, конечностей. Гемолимфа представляет бесцветную или желтоватую жидкость, у немногих насекомых содержит гемоглобин. Клетки крови – гемоциты, их функции разнообразные.
Плазма содержит аминокислоты, растворенные соли, углеводы, мочевую кислоту и другие вещества. Функции гемолимфы: транспорт питательных веществ, защитная, гидравлическая, метаболизм, отчасти дыхательная.
91. Выделительная и половая системы насекомых. Метаморфоз и его основные типы. Выделительная система: Функции органов выделения у насекомых выполняются рядом образований. В первую очередь, это мальпигиевы сосуды. Они эктодермального происхождения, располагаются на границе между средней и задней кишкой. Их полость выстлана однослойным эпителием. Количество мальпигиевых
сосудов колеблется от 2 (червецы) до 150 (пчелиные). Открываются сосуды самостоятельно либо объединены в пучки. Мальпигиевы сосуды обладают некоторой подвижностью, что обеспечивает постоянную смену омывающей их гемолимфы. Из гемолимфы продукты обмена в виде растворенных в воде солей мочевой кислоты поступают в полость сосудов, где образуется мочевая кислота. Освобождающаяся при этом вода всасывается стенками сосудов и поступает обратно в гемолимфу. Особенно интенсивно эти процессы идут в задней кишке, куда поступают продукты обмена из мальпигиевых сосудов. Ректальные железы являются основным местом всасывания воды. Почти сухие кристаллы мочевой кислоты вместе с экскрементами выводятся через анальное отверстие. Подобное выведение продуктов обмена в виде сухих кристаллов позволяет насекомым экономно расходовать воду, поступающую в организм. У форм, обитающих в среде с повышенной влажностью или поглощающих большое количество жидкой пищи, реабсорбции воды не происходит. Кроме мальпигиевых сосудов, функцию выделения выполняют лабиальные железы. Они представляют собой парные образования, открываются одним отверстием в основании нижней губы. Лабиальные железы характерны насекомым отрядов подуры, ногохвостки, щетинохвостки. Вторая группа органов выделения – органы накопительной экскреции. Ими являются жировое тело (уратные клетки), нефроциты и у некоторых (тизануры, прямокрылые) фагоцитарные органы Половая система насекомых: Насекомые раздельнополые (за редким исключением) животные, нередко обладают резко выраженным половым диморфизмом. Половые железы парные. У самок каждый яичник состоит из яйцевых трубок – овариол, в начальном участке – гермарии, образуются яйцеклетки. Овариолы крепятся в организме с помощью концевого филамента. Овариола заканчивается ножкой, совокупность ножек образует каликс, продолжающийся в яйцевод. Оба
яйцевода сливаются в непарное влагалище, открывающееся на брюшной стороне брюшка. Во влагалище открывается семяприемник, нередко и совокупительная сумка, придаточные железы. В области полового отверстия могут находиться яйцеклады. Мужская половая система состоит из пары семенников, каждый из которых представлен трубочками – фолликулами. Семяпроводы объединяются в общий семяизвергательный канал, в последний часто открываются дополнительные железы. Семяизвергательный канал пронизывает совокупительный орган, у насекомых, не имеющих такового, живчики склеиваются в сперматофоры, одетые оболочкой. Метаморфоз:Первичные формы метаморфоза характерны для низших насекомых. Выделяют 2 типа: анаморфоз (отряд протуры) и протоморфоз (у насекомых отрядов двухвостки, коллемболы, щетинохвостки). Основные формы метаморфоза: гемиметаболическое и голометаболическое развитие (превращение). Первая форма характеризуется прохождением следующих фаз развития: яйцо (ova), личинка (larva), взрослое насекомое (imago). Личинка похожа на взрослое насекомое (нимфа), ведет сходный с ним образ жизни (таракановые, полужесткокрылые и др.) или не похожа на взрослое насекомое (наяда), имеет провизорные органы (стрекозы). Разновидностями гемиметаморфоза являются гипоморфоз и гиперморфоз. При голометаболическом развитии насекомое проходит 4 стадии: яйцо, личинка, куколка (pupa), взрослое животное. Такое развитие характерно насекомым отрядов перепончатокрылые, двукрылые, жесткокрылые, чешуекрылые и др. Личинка при данном типе метаморфоза не похожа на взрослое животное и ведет иной образ жизни. Процесс метаморфоза контролируется железами внутренней секреции. В процессе метаморфоза существенное значение имеет деятельность прилежащих тел, продуцирующих ювенильный гормон. При высокой его концентрации линька личинки приводит к образованию личинки следующего возраста. Разновидностью голометаморфоза является гиперметаморфоз. Он характерен нарывникам, веерокрылым и некоторым другим насекомым. Происхождение метаморфоза. Существует несколько гипотез о происхождении метаморфоза. Согласно одной из них обе формы метаморфоза возникли независимо друг от друга от более простого типа развития – протоморфоза. Другая предполагает постепенное происхождение метаморфоза:
протоморфоз – гемиметаморфоз – голометаморфоз. Последнее время все больше исследователей склоняются к первой гипотезе.
92. Характеристика классов Protura, Collembola, Diplura. Бессяжковые или протуры (лат. Protura, от греч. protos — «первый» и oura — «хвост») — отряд скрыточелюстных шестиногих. Более 500 видов, в том числе около 90 в Европе и в России. Малоизученная группа, по некоторым оценкам, истинное число видов превышает число описанных более чем в 1,5 раза. Строение: Тело удлиненное, 0,5 — 2 мм в длину. Ротовой аппарат энтогнатного типа, специализированный, мандибулы колющие. Протурам свойственен ряд уникальных апоморфий и плезиоморфий в строении тела. Голова лишена каких-либо следов глаз и, что особенно уникально среди членистоногих, антенн. Имеются постантеннальные органы — ложные глазки. Передние ноги сильно вытянуты и снабжены большим количеством сенсоров, заменяя утраченные усики. Строение груди и первых сегментов брюшка различаются слабо. На последних присутствуют двухчлениковые слаборазвитые ноги либо их одночлениковые рудименты, сохранившие мускулатуру. Лапка одночлениковая, свободная. VIII сегмент брюшка модифицирован. На заднем крае VIII тергита брюшка открывается пара абдоминальных желез, причем каждое отверстие замкнуто изнутри кутикулярной пробочкой, от которой к передне-латеральной части тергита тянется мускул-ретрактор. Между VIII и последним сегментом имеется еще три вторичных сегмента меньшего размера, развивающихся в ходе онтогенеза (то есть количество сегментов увеличивается от 9 у личинки I возраста до 12 у личинки III возраста). У некоторых бессяжковых сохраняется рудиментарная примитивная трахейная система. Развитие и образ жизни: Появление дополнительных сегментов брюшка в процессе развития дало предполагать наличие у бессяжковых анаморфоза и на этом основании сближать их с многоножками, противопоставляя остальным насекомым. Однако существует и другое объяснение природы этих сегментов. Согласно ему, IX—XI сегменты — ложные, образовавшиеся в результате вторичного деления VIII сегмента, и поэтому они резко отличаются от истинных сегментов размером и способом развития в онтогенезе. В пользу этого говорит строение нервной системы: от абдоминальных ганглиев отходит лишь 9 пар сегментальных нервов, как и у других насекомых. При такой интерпретации, половое отверстие оказывается расположенным не между XI и XII, а между VIII и IX сегментами, то есть как у двухвосток. Кроме увеличения числа видимых сегментов, метаморфоз протур интересен наличием у самца стадией преимаго с недоразвитыми гениталиями, за которой следует половозрелая стадия имаго. Линьки половозрелых особей не наблюдались. Бессяжковые — обитатели почвы и разлагающихся растительных останков. Удельное количество бессяжковых на 1 квадратный метр земли составляет около 90000. Большинство питаются грибами, высасывая их мицелий, некоторые являются хищниками. Между предпоследним и последним сегментами брюшка у самки и самца имеются выпячивающиеся гениталии, состоящие из непарной части (перифаллус у самца и перегина у самки), несущей спереди пару уходящих вглубь тела базальных аподем, а сзади — пару двучлениковых придатков. Семенные протоки самцов на всем протяжении парные, открываются на дистальных члениках придатков. Влагалище самки непарное, открывается между основаниями придатков. Collembola:Коллемболы или ногохвостки (Collembola, англ. Springtail) — отдельный отряд членистоногих (около 10 тыс. видов). Ранее рассматривался как отряд первично бескрылых насекомых, однако в современной классификации отнесён к членистоногим, иногда рассматривается в ранге класса. Это шестиногие микроартроподы размером 0,1-9 мм, населяют, главным образом, подстилку и почву во всех регионах Земли. По внешнему виду чётко разделяются на вытянутых, иногда червеобразных членистобрюхих и шарообразных слитнобрюхих. Также довольно многочисленны и в других местообитаниях, таких как стволы деревьев, лишайники. Характерных представителей можно в массе повстречать на поверхности водоёмов (Podura aquatica), в траве (Sminthuridae), на снегу в горах (Entomobrya nivalis). Отдельно стоит сказать о коллемболах, заселяющих приливно-отливную зону на морском берегу, так как у них можно наблюдать ряд интересных приспособлений для жизни в данном местообитании. Считается, что ногохвостки возникли в девоне (ок. 420 млн лет назад), откуда известны их первые палеонтологические находки (Rhyniella praecusor). В настоящее время представляют интерес как для фундаментальной науки, так и для практического применения, являясь биоиндикаторами при выявлении загрязненности почв. ДВУХВОСТКИ (DIPLURA) Отряд объединяет около 300 видов, обитающих в почве, в муравейниках и гнилой древесине. Такое название животные получили благодаря парным придатках, расположенным на заднем конце тела. Глаза отсутствуют. Обособленная голова несёт длинные усики. Направленные вверх ротовые органы скрыты в головной капсуле. Ноги длинные. Брюшко удлинённое Представители семейства япиксов (Japygidae) - небольшие двухвостки около 1 см, обитающие в жарком поясе. На заднем конце тела эти двухвостки имеют твёрдые клешневидные придатки, с помощью которых, как клешнями, удерживают добычу.
93. Открыточелюстные или безмышцеусые (лат. Ectognatha или Amyocerata) Открыточелюстные или безмышцеусые (лат. Ectognatha или Amyocerata) — подкласс насекомых, характеризующийся «обычным» строением ротового аппарата, в отличие от скрыточелюстных. К открыточелюстным относится подавляющее большинство насекомых. Иногда только открыточелюстных называют насекомыми (одновременно перенося на них латинское название Insecta), но такая точка зрения не имеет под собой серьезных оснований. Общие признаки: Открыточелюстных объединяют следующие признаки: 1. В антенне только два членика приводятся в движение мышцами: рукоятка приводится в движение мышцами, находящимися в голове, и содержит в себе мыщцы, приводящие в движение второй членик — ножку. Внутри ножки располагается джонстонов орган — слуховой орган, у которого сколопсы прикреплены к сочленовой мембране между рукояткой и ножкой. От ножки отходит также вторично расчлененный жгутик, у наиболее примитивных открыточелюстных состоящий из большого неопределенного числа члеников и имеющий щетинковидную форму. Такая форма жгутика связана с тем, что в процессе онтогенеза число члеников жгутика увеличивается за счет деления пополам члеников в проксимальной части жгутика и последующему их удлинению. У многих крылатых насекомых число вторичных члеников стабилизируется и жгутик приобретает иную форму (булавовидную, веретеновидную и др.); 2. Имеется 3 теменных простых глазка. Эти глазки, в отличие от сложных глаз, иннервируются не от зрительных долей мозга, а от особого глазкового центра. Они гомологичны науплиальным глазам ракообразных, некоторым фоторецепторам хелицеровых и, предположительно, теменным глазкам трилобитов, так что имеют древнее происхождение. Особенностью же глазков открыточелюстных насекомых является то, что передний из них непарный, образовавшийся в результате слияния пары глазков. Функция их до конца не выяснена, неясно также, почему они подверглись редукции у многоножек и скрыточелюстных насекомых (доказательством исходного наличия двух пар теменных глазков у последних служит обнаружение их внутренних гомологов у ногохвосток); 3. Имеется тенториум (tentorium) — эндоскелет головы, имеющий эктодермальное происхождение. В состав тенториума входит пара передних рук и задний тенториальный мост. Передние руки тенториума — это апофизы, начинающиеся от переднего края головной капсулы около передних мыщелков мандибул. На внешней поверхности головы заметна пара углублений — передних тенториальных ямок, ведущих в передние руки тенториума. Снаружи эти ямки нередко соединены поперечной бороздкой, разделяющей лоб и наличник. Задний тенториальный мост образован парой слитых между собой апофизов, начинающихся в задней части головы между основаниями максилл и нижней губы и сливающихся между собой под пищеводом, над вентральной нервной цепочкой и позади окологлоточных коннектив. Наружние углубления, ведущие в задний мост, называются задними тенториальными ямками. По мнению некоторых авторов, передние руки тенториума гомологичны фольземовым рукам ногохвосток и аподемам фультур многоножек; 4. Лапка вторично расчленена на 5 члеников (тарзитов), не имеющих собственных мышц. У многих открыточелюстных происходит вторичное уменьшение числа члеников вплоть до полной утраты расчлененности, но никогда не бывает больше 5 члеников. В других группах неполноусых лапки либо нерасчлененные, либо расчленены на несколько члеников (всегда меньше 5); 5. У самок имеется яйцеклад (oviductus), образованный сложенными вместе гонапофизами (gonapophyses) — своеобразными парными вентральными придатками VIII и IX сегментов брюшка, называемыми также первыми и вторыми створками яйцеклада. Гонапофизы представляют собой длинные выросты, отходящие от углов кокситов. Существует предположение, что они являются видоизмененными остатками конечностей своих сегментов. Благодаря тому, что IX сегмент брюшка у самки практически полностью редуцирован, и его кокситы почти до основания вжвинуты под кокситы VIII сегмента, основания обоих пар гонапофизов оказываются вплотную прижатыми друг к другу, образуя яйцеклад. У разных групп крылатых насекомых яйцеклад редуцируется, а в качестве функциональной замены ему образуется вторичный яйцеклад различных типов; 6. Половое отверстие самца открывается на границе IX и X сегментов брюшка, а не VIII и IX, как у скрыточелюстных. Обычно половое отверстие открывается на пенисе, который представляет собой вырост в сочленении IX стернита с X сегментом. Интересно, что у щетинохвостковых пенис, хотя и имеется, но никогда не вводится в половые пути самки, поэтому некоторые исследователи полагают, что его исходная функция не была связана с копуляцией; 7. На нижней стороне X сегмента вместо непарного стернита имеется пара парапроктов (paraprocti), а позади X тергита имеется эпипрокт (epiproctus), считающийся рудиментом XI сегмента брюшка; кроме того, конец брюшка, помимо церков, несет непарный парацерк, имеющий аналогичное церкам строение и в большинстве высокоорганизованных групп открыточелюстных вторично утраченный. Существуют гипотезы об исходном наличие парацерка у двухвосток, поэтому апоморфный статус парацерка спорен. Неясно и его происхождение; 8. Сегментарные дыхальца груди утрачены (в отличие от двухвосток и бессяжковых). Классификация: Существуют несколько вариантов классификации открыточелюстных. В двух из них щетинохвостковые не признаются как голофилетический таксон, и одна из групп щетинохвостковых предлагается в качестве сестринской к крылатым насекомым. Наиболее распространенный вариант предполагает деление открыточелюстных на два инфракласса: Triplura и Pterygota. В классификациях, основанных на гипотезе о полифилии крылатых насекомых, открыточелюстные делятся и на большее число равноправных таксонов. С выделением скрыточелюстных в равноправный класс наравне с насекомыми в составе надкласса шестиногих, таксон открыточелюстных стал тождественен насекомым.
94. Triplura. Щетинохвостки (лат. Thysanura или лат. Zygentoma) — отряд первичнобескрылых насекомых. Одна из наиболее характерных особенностей — наличие трёх хвостовых нитей (пары церков и непарного парацерка), благодаря которым щетинохвостки и получили своё название. Из всех первичнобескрылых щетинохвостки наиболее близко родственны крылатым насекомым, с которыми их сближает строение усиков, или антенн, и сочленения верхних челюстей, или мандибул, с головной капсулой. Вместе с тем, щетинохвостки характеризуются рядом примитивных особенностей: длинными членистыми церками, наличием парацерка, рудиментарными конечностями на брюшных сегментах (втяжными пузырьками и грифельками). Наиболее известный представитель — обыкновенная чешуйница, нередко встречающаяся в жилых домах. В мировой фауне более 400 видов, в основном обитающих в тропических и субтропических поясах. Щетинохвосток часто ошибочно называют мокрицами.
95. Крылатые насекомые (лат. Pterygota) — инфрокласс насекомых, в котором объединены все виды, оснащённые крыльями, включая тех, которые в ходе эволюции вновь утратили способность к полёту (к примеру, блохи или вши). К крылатым насекомым относится бо́льшая часть всех насекомых за исключением первичнобескрылых щетинохвосток (Zygentoma) и древнечелюстных (Archaeognatha). С развитием у насекомых крыльев был связан также ряд морфологических и физиологических изменений, из-за которых можно с уверенностью делать вывод, что процесс в эволюции насекомых, приведший к способности насекомых летать, произошёл всего один раз и все крылатые насекомые являются потомками одного общего вида. Крылья насекомых представляют собой тонкую хитиновую пластинку, пронизанную трахеями. Вероятно, они развились из боковых пластинок (Paranota) на всех трёх сегментах торакса. У современных крылатых насекомых остались, однако, только две пары крыльев, расположенные на втором и третьем сегменте торакса. Третья пара встречается у ископаемых видов, таких как Stenodictya. Трахеи для снабжения крыльев ответвляются от трахей конечностей. Так как крылья развились в качестве удвоения телесной оболочки, то трахеи расположены внутри этих двух слоёв. С тораксом крылья соединены с помощью сложного сустава, который у стрекоз напрямую связан с мышцами. Намного более распространённым вариантом является взмах крыльев посредством расширения и сжатия грудного сегмента.
96. Прямокрылые - одна из наиболее известных групп насекомых не только потому, что все знают крупных кузнечиков и сверчков, но и потому, что среди них очень много опасных вредителей сельского хозяйства. Издавна и до сих пор полчища саранчи уничтожают огромные площади посевов, оставляя за собой голую землю. Огородникам хорошо знакома медведка. Большинство прямокрылых - крупные насекомые, обычно с хорошо развитыми крыльями, сильными задними ногами и крупной головой с грызущими челюстями. Многие прямокрылые питаются растительной пищей (большинство саранчовых), часть видов всеядна (кузнечики, сверчки и медведки), среди кузнечиков встречаются и хищники. Усики могут быть длинными (длиннее тела) и более короткими (короче половины тела). По этому признаку прямокрылых разделяют на 2 подотряда - длинноусых (Dolichocera) и короткоусых (Brachycera). Прямокрылые - насекомые с неполным превращением. Из яиц у них выходят личинки, которые имеют такой же вид, как и взрослые насекомые, только очень маленькие. Кроме того, у личинок не развиты крылья. В остальном они точная копия своих родителей и питаются так же, как и взрослые насекомые. У прямокрылых хорошо развиты органы зрения (два крупных сложных глаза и 3 маленьких глазка) и органы слуха; они могут издавать и воспринимать звуки при помощи различных аппаратов. Звуковой аппарат у кузнечиков (надсемейство Tettigonioidea) располагается на передних крыльях: во время стрекотания кузнечик слегка приподнимает и раздвигает крылья, а затем начинает вибрировать ими так, что зубчики на одном крыле трутся о толстую жилку на другом. Каждый вид кузнечиков издает характерную только для него песню, по которой можно определить вид певца. "Пение" наших обычных певчих кузнечиков, широко распространенных повсеместно в лесной зоне, можно наблюдать в садах и на опушках, где кузнечики сидят на ветвях кустов. Найти певца можно только по звуку, тогда как увидеть кузнечиков очень трудно, потому что все они имеют покровительственную окраску под цвет растений, на которых живут. Песня была бы бесполезна, если бы кузнечики не имели органа слуха. Слуховые аппараты расположены у них на голенях передних ног, это два щелевидных отверстия, затянутых тонкой плёнкой, выполняющей роль барабанной перепонки. Звуковой аппарат сверчков (надсемейство Grylloidea) устроен так же, как у кузнечиков. Стрекочут у сверчков только самцы, при этом их песни отличаются не только у разных видов, но и у одного и того же сверчка в разных ситуациях. Саранчовые (надсемейство Acridoidea), относящиеся к подотряду короткоусых, также признанные музыканты, однако их органы стрекотания и слуха устроены по-другому. Саранчовые издают звуки, быстро потирая зубчатыми бёдрами задних ног по жилке на крыле. Некоторые саранчовые, например трещотки, издают резкие трескучие звуки во время полёта. Органы слуха у саранчовых расположены по бокам брюшка, на первом брюшном сегменте. Также как и у кузнечиков, они представляют собой щели, затянутые перепонками. Прямокрылые составляют довольно многочисленный отряд насекомых, насчитывающий свыше 20000 видов, из которых более 700 встречаются на территории России и прилежащих стран.
97. Полужесткокрылые, хоботные или клопы (лат. Hemiptera s. Rhynchota) — отряд насекомых. Из двух названий более удачным следует считать второе, так как весьма употребительное первое название указывает на особенности, свойственные лишь одному из подотрядов. К наиболее характерным признакам отряда относятся ротовые органы в виде членистого хоботка и неполное превращение. Отряд КЛОПЫ представляет собой самый крупный отряд насекомых с неполным превращением. В настоящее время известно свыше 30 000 видов клопов. Свое название "полужесткокрылые" они получили благодаря своеобразному строению передних крыльев, резко отличающихся от задних, имеющих вид прозрачных перепонок с небольшим количеством жилок. Передние крылья превращены в надкрылья, неоднородные по степени их хитинизации. Основная часть надкрылья состоит из твердого хитина, в то время как вершинная часть перепончатая и жилки на ней хорошо заметны. Некоторым видам клопов свойствен так называемый крыловой диморфизм, сущность которого заключается в том, что наряду с полнокрылыми особями с вполне развитыми крыльями и надкрыльями у них имеются и короткокрылые особи Помимо структуры надкрылий, характерным признаком, свойственным всем представителям этого отряда, является их колюще-сосущий ротовой аппарат, имеющий вид хоботка, отходящего от переднего края головы и не срастающегося с переднегрудью. Основную часть хоботка представляет сильно вытянутая членистая нижняя губа, образующая на внутренней стороне глубокий желобок, в котором помещаются сильно измененные верхние и нижние челюсти, имеющие вид тонких и длинных щетинок. Сверху хоботок прикрывается относительно недлинной верхней губой. В связи с такой специфичной структурой ротового аппарата находится характер питания полужесткокрылых: прокалывая покровы тела животного или растения, клопы высасывают кровь или клеточный сок. У растительноядных клопов хоботок обычно длинный и тонкий, в спокойном состоянии он подогнут под тело и скрыт в особом желобке, расположенном на голове и груди; у хищных видов хоботок, наоборот, короткий, но толстый и сильный, дугообразно изогнутый в виде клюва. Число личиночных стадий у полужесткокрылых в преобладающем большинстве случаев пять, редко четыре. Наиболее древние находки насекомых, идентичных современному отряду полужесткокрылых, относятся к мезозойской эре, где они в основном были представлены специфическими семействами, не встречающимися позднее ни в третичной, ни в современной фауне. В третичном периоде, по крайней мере уже с нижнего олигоцена, обособляется большинство современных семейств клопов. В настоящее время насчитывают приблизительно 50 семейств, из которых около 40 (свыше 2000 видов) встречаются в России. Преобладающее же большинство видов населяет преимущественно тропические и субтропические области земного шара. Среди клопов много видов хищных или со смешанным питанием, но преобладают растительноядные формы. Некоторые из них яляются вредителями сельскохозяйственных лесных растений. Многие клещи (особенно из родов Orius и Nabis) полезны, т.к. уничтожают вредных для сельского и лесного хозяйства тлей, клещей, гусениц, личинок жуков и др..
98.Жесткокрылые или Жуки.(Coleoptera) — Ж. составляют один из наиболее характерных отрядов насекомых. Отличительные признаки их: более или менее твердые крылья первой пары, служащие лишь крышкой для задних, жевательные органы рта и полное превращение. Как форма тела, так и величина крайне разнообразны. Голова редко свободна, обыкновенно несколько вдается в 1-й членик груди. Усики или сяжки очень разнообразной формы и величины, обыкновенно из 11 члеников (их может быть от 4 до 30 и более). Сложные глаза умеренно развиты, иногда каждый подразделяется хитиновым гребнем на два. Редко (например, живущие в пещерах или муравейниках) слепы. Глазков по большей части нет, иногда же бывает 1-2. Органы рта типические жевательные, за исключением Nemognatha из семейства шпанских мух (Cantharidae), у которого нижние челюсти вытянуты, как у бабочек, в хоботок, челюстные щупики 4-члениковые, губные — 3-члениковые. Из грудных колец наиболее развито и подвижно первое, образующее характерный для Ж. грудной щит. Среднее развито меньше всего и на спинной стороне несет треугольную пластинку — щиток, вдающийся между основаниями надкрылий; заднее развито больше. Передние крылья Ж. (надкрылья или элитры) более или менее тверды и кожисты, часто покрыты различными возвышениями и углублениями и служат лишь покрышкой для задних крыльев; обыкновенно плотно прилегают друг к другу по средней линии, образуя так называемый шов, по которому иногда и срастаются между собой; редко надкрылья расходятся и отчасти налегают друг на друга. Надкрылья или покрывают сверху все брюшко, или оставляют непокрытой заднюю часть — pygidium, или укорочены и прикрывают лишь небольшую часть брюшка. Нижние крылья, служащие для летания, в покое складываются по большей части не только вдоль, но и (раз или два) поперек; иногда их вовсе нет. Полет большей части Ж. довольно тяжел. Ноги обыкновенно бегательные, но иногда плавательные (причем ноги задней пары уплощенны, расширены и усажены щетинками), роющие (особенно передняя пара, голень которой в таком случае расширена и зубчата), прыгательные и т. д. Лапки обыкновенно с 5, реже 4 или 3-1 члениками. Брюшко прикреплено к груди широким основанием и по большей части (кроме форм с укороченными элитрами) покрыто сверху мягкой кожей; на верхней стороне его заметно больше колец (обыкновенно 8), чем на нижней (4-7); задние членики втянуты внутрь брюшка и преобразованы в наружные половые органы. Нервная система представляет от 2 до 13 узлов. Дыхательные органы часто с расширениями. Покровы Ж. часто очень тверды и иногда отличаются чрезвычайно яркими и блестящими цветами. Некоторые представляют резкий миметизм. Лишь немногие Ж. живородящи, некоторые кладут яйца в особое гнездо. Превращение полное. Личинки всегда снабжены хорошо развитой головой, 6-ногие или безногие, чаще всего беловатого или желтоватого цвета в связи с жизнью в темноте (в растениях, под землей, в гниющих веществах и т. п.); свободно живущие черного, реже — других цветов. Некоторые Ж. представляют гиперметаморфоз. Куколка Ж. неподвижна, все части взрослого животного ясно различимы и свободны (pupa libera); кокона обыкновенно нет, но некоторые, живущие в дереве, делают кокон из частичек дерева, у некоторых долгоносиков встречаются настоящие коконы, сплетенные из нитей. Некоторые Ж. приносят прямую пользу человеку: так, личинки пальмового долгоносика и некоторых других употребляются в пищу, шпанская мушка (Lytta vesicatoria) употребляется в медицине, наконец, различные красиво окрашенные или светящиеся Ж. употребляются как украшения. Несравненно большее значение имеет косвенная польза многих Ж.; именно хищные формы, питаясь вредными насекомыми, приносят большую пользу земледелию, садоводству и лесоводству. Многие виды приносят и большой вред лесам и полям, другие поедают запасы пищи (например, муку, зерновой хлеб, плоды и т. д.), меха, кожу и т. д.
99. Сетчатокрылые (лат. Neuroptera). Сетчатокрылые (лат. Neuroptera, синоним: Planipennia) — отряд свободноживущих новокрылых насекомых с полным превращением. Эта относительно небольшая группа включает от 3,5 до 5 тысяч видов. Сетчатокрылые имеют вытянутое тело с мягкими покровами. 2 пары крыльев этих насекомых покрыты густой сетью жилок. Окраска сетчатокрылых нежно-зеленая или бурая, зачастую с ярко-золотистыми глазами. К отряду относятся такие представители, как златоглазки, муравьиные львы, мантиспы. Сетчатокрылые — преимущественно хищные насекомые. Наибольшие отличия в морфологии и экологии представителей отряда наблюдаются на личиночной стадии. Впервые сетчатокрылые появились в Пермском периоде. Дальнейшему развитию группы способствовали климатические и геологические изменения Мезозойской эры.Общая характеристика Взрослые особи сетчатокрылых имеют длину тела 2—20 мм, крылья в размахе достикают 120 мм. К крупным представителям отряда относятся, например, отдельные виды муравьиных львов (Myrmeleontidae), а также некоторые аскалафусы (Ascalaphidae). Голова сетчатокрылых гипогнатическая, со сложными глазами, между которыми расположены усики. Ротовой аппарат грызущий. У личинок ротовой аппарат функционирует как колюще-сосущий. Антенны сетчатокрылых многочленистые, иногда головчатые. Отличительной особенностью сетчатокрылых является наличие у взрослых особей двух пар развитых сетчатых крыльев, складывающихся крышеобразно. Крылья обычно прозрачные, пятнистые, но могут быть окрашенными. У большинства видов сеть жилок на крыльях богатая, реже упрощенная, при этом ветви у края заметно раздвоены. Мускулатура крыльев сетчатокрылых полностью или частично гомономная. В полете передняя и задняя пары крыльев двигаются независимо. Сетчатокрылые — насекомые с полным превращением. Это означает, что личиночная и имагиальная стадии сильно различаются по экологическим и морфологическим характеристикам. Куколки сетчатокрылых свободные, иногда имеют волокнистый кокон, который формируется за счет секрета мальпигиевых сосудов, открывающихся в задний отдел кишечника.Пищеварение у личинок сетчатокрылых наружное Пищеварительный секрет впрыскивается в тело жертвы по каналу, образованному желобком мандибул и прилегающей к нему внутренней жевательной лопастью нижних челюстей. По этому же каналу всасывается разжиженное пищеварительными соками внутреннее содержимое жертвы. Причем у личинок сетчатокрылых непереваренные остатки пищи не выводятся наружу, а накапливаются в течение всего развития. Это обеспечивается наличием непроницаемой перегородки между средней и задней кишкой личинок. При образовании имаго происходит соединение средней и задней кишки, и накопленные ранее экскременты выбрасываются наружу. Для ювенильных особей всех сетчатокрылок характерно наличие трех пар ног и отсутствие нижнечелюстных щупиков. При этом ноги личинок заканчиваются пятичлениковыми лапками с двумя коготками Сетчатокрылые значительно различаются не только по внешнему виду, но и по организации жизненного цикла. Для полноты картины рассмотрим развитие нескольких основных семейств отряда.
100. Филогения членистоногих. Членистоногие по своему происхождению связаны с кольчатыми чер-
вями. В процессе филогенеза возникают конечности на каждом сегменте, обосабливается головной отдел, конечности головы специализируются на выполнении функций ротового аппарата и превращаются в таковой. Сегментация первоначально гомономная, в последующем у большинства представителей модифицируется в гетерономную. Покровы: склеротизация покровов, образование внешнего скелета из тонкой кутикулы червей. Образование членистой конечности. Мышцы: распад кожно-мускульного мешка на отдельные мышцы. Появление смешанной полости тела (миксоцеля), обособление сердца, развитие сложных глаз. Наиболее примитивные формы встречаются в подтипах трилобитообразных и ракообразных. Мандибулятные по ряду признаков схожи с ракообразными, которых производят от трилобит. Хелицеро-
вые по своему происхождению связаны с трилобитами. Существуют и другие гипотезы филогении членистоногих: все подтипы типа произошли от разных предков первичных членистоногих.
101.Подотдел Lophophorata. Особенности строения. Щупальцевые составляют группу вторичнополостных животных, олигомерных (малосегментных) животных не вполне выясненного происхождения. Щупальцевые ведут сидячий образ жизни, который оказал глубокое влияние на их организацию. Характерные признаки типа: 1. Тело неясно расчленено и слагается из трёх сегментов. Первый-предротовая лопасть, или эпистом.Второй сегмент несёт ротовое отверстие. Он окружён ресничными щупальцами, которые служат для подгона пищевых цастиц ко рту и для дыхания. Щупальца сидят на щупальценосце, или лофофоре, в основе имеющем форму подковы, или расположены по кругу. Третий сегмент собственно туловищный. В ряде случаев эпистомальный сегмент вторично редуцируется. 2. Полость тела представлена целомом, который в соответствии с сегментацией разделяется на три отдела:ипередний эпистомальный, средний-сравнительно узкий, посылающий каналы в лофофор и щупальца, и задний, самый обширный – туловищный целом. Обособленность отделов целома может быть не всегда хорошо выражена. 3. Анальное отверстие лежит недалеко от рта, кишечник образует петлю. 4. Имеется кровеносная система, которая может вторично редуцироваться. 5. Органы выделения (нефридии) представлены одной или двумя парами ресничных воронок. Открывающихся в целом. Являются ли эти образования метанефридиями или целомодуктами, пока окончательно неустановленно. Иногда органы выделения редуцированы.6. Половые железы образуются в туловищном целоме. 7. Дробление яйца полное, не спирального типа, из яйца развивается планктонная личинка, более или менее напоминающая трохофору.
102. Класс мшанок. Тип вторичноротых беспозвоночных, представленный прикрепленными колониальными формами. В палеозое создавали рифовые постройк. Класс мшанок содержит водных, преимущественно морских, реже пресноводных животных, ведущих сидячий образ жизни и образующих в результате бесполого размножения колонии. Чаще всего колонии бывают древовидными, ветвистыми. В некоторых случаях ветви колонии сплющиваются в одной плоскости и принимают характер пластинок, а сама колония становится похожей на пучок листьев. Если же колонии стелются по субстрату, то образуют на дне или на стеблях водорослей тонкие корочки. Каждая колония объединяет большое количество особей. Таким образом, по внешнему виду колонии мшанок больше всего напоминают гидроидных полипов. Размеры мшанок невелики: колонии измеряются несколькими сантиметрами, отдельные особи большей частью длиной не более 1 мм. Современных видов мшанок около 4000, ископаемых еще больше. Мшанки живут преимущественно в море, где встречаются на различных глубинах, начиная с прибрежных камней и до больших глубин. В пресных водах встречаются почти исключительно снабженные щупальценосцем Phylactolaemata. Наиболее обыкновенны из них виды рода Plumatella, образующие стелющиеся по субстрату, ветвящиеся колонии или массивные наросты на подводных предметах, и Cristatella mucedo. Последняя особенно интересна тем, что у нее проявляется высокая степень интеграции колонии, выражающаяся в слиянии цистидов. Колонии Cristatella имеют вид студенистой колбаски, на верхней поверхности которой торчат полипиды. Подошва не прикреплена к субстрату, и вся колония может очень медленно ползать. Срок жизни пресноводных колоний обычно не больше 5-6 месяцев; колонии редко перезимовывают. Жизнь отдельных особей колоний значительно короче; на колонии всегда можно встретить дегенерирующие особи, места которых впоследствии занимаются индивидами, вновь развивающимися из почек. Древнейшие морские мшанки известны уже из силурийских отложений. Практическое значение мшанок мало. Наряду с другими беспозвоночными животными мшанки составляют важный компонент подводных обрастаний. Кроме того, в водопроводах, не снабженных хорошими песчаными фильтрами, мшанки могут причинять существенней вред, закупоривая трубы и давая приют разным мелким организмам, загрязняющим воду. Отдельные особи мшанок подразделяются на два отдела, которые, однако, не являются сегментами. Передний конец особи свободно торчит в окружающую воду и несет на себе рот с венчиком щупалец вокруг. Задняя половина животного одета утолщенной кутикулой, производимой наружным эпителием тела, и имеет вид глубокой чашечки или мешочка. Эту часть животного нередко обозначают особым термином - цистид. Передняя половина мшанки лишена плотной кутикулы, нежная и при раздражении втягивается внутрь цистида; ее называют полипидом. Втягивание совершается при помощи двух особых мускулов-ретракторов. Они находятся по бокам кишечника и передним концом прикрепляются изнутри к стенке тела полипида по сторонам рта, а задним концом - к основанию цистида. При втягивании передней половины тела в цистид отдельные щупальца не вворачиваются, а весь венчик их уходит в глубь образующегося впячивания. Щупальца имеют вид длинных нежных выростов, в которые заходят каналы целома, и покрыты мерцательным эпителием; они служат для собирания пищевых частиц и в качестве органов дыхания. У пресноводных мшанок, образующих подкласс Phylactolaemata (Покрыторотые), щупальца в два ряда располагаются на особом щупальценосце - лофофоре, имеющем сверху, со стороны рта, форму подковы, у вершины которой и располагается ротовое отверстие. Последнее прикрыто небольшим язычковидным выростом - эпистомом - предротовым отделом тела. Представители подкласса Gymnolaemata вторично утратили эпистом, а их щупальце вокруг ротового отверстия образуют простой венчик. Кутикула цистида может быть различной консистенции: в виде тонкой хитиновой пленки или остуденяется и превращается в толстый желеобразный слой, содержащий до 99% воды, В других случаях кутикула утолщается и пропитывается углекислой известью, становясь очень твердой. Наружный эпителий мшанок однослоен и у пресноводных форм (подкл. Phylactolaemata) подстилается двумя слабо развитыми слоями мышц: наружным из кольцевых и внутренним из продольных мышечных волокон. У морских голоротых мшанок (подкл. Gymnolaemata) мышечные слои отсутствуют. Слабое развитие кожно-мускульного мешка объясняется неподвижностью мшанок. Перитонеальный эпителий ограничивает обширную вторичную полость тела, в которой помещаются все внутренности. Полость тела (целом) тонкими перегородками делится на три отдела. Передний обладает небольшими размерами и залегает в эпистоме. Средний - кольцевой канал - окружает глотку и посылает слепые ответвления в щупальца. Задний, наиболее обширный, занимает почти все тело и называется туловищным целомом. У голоротых мшанок эпистом вместе с целомом редуцируется.
|
|
|
|
|
Дата добавления: 2015-04-24; Просмотров: 723; Нарушение авторских прав?; Мы поможем в написании вашей работы!