КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Роль мембран и их разнообразие
|
|
|
|
Мембраны играют ключевую роль как в структурной организации, так и в функционировании всех клеток - прокариотических и эукариотических, растительных и животных. Мембраны формируют внутриклеточные компартменты, с их помощью происходит разделение содержимого компартментов и окружающей их среды. Но если бы это была единственная функция мембран, они не были бы для нас столь интересны. Мембраны не только разделяют клетку на отдельные компартменты, но и участвуют в регуляции всех связей и взаимодействий, которые осуществляются между наружной и внутренней сторонами этих компартментов. Это может проявляться в виде физического переноса ионов или молекул через мембрану или в форме передачи информации при помощи конформационных изменений, индуцируемых в мембранных компонентах. Кроме того, с мембранами связаны многие клеточные ферменты. Некоторые из них катализируют трансмембранные реакции, когда реагенты находятся по разные стороны мембраны или когда каталитический акт сопровождается транспортом молекул. Другие ферменты образуют своеобразные комплексы, которые осуществляют цепь последовательных превращений, причем благодаря тому, что эти ферменты располагаются в плоскости мембраны, повышается эффективность всего процесса. Имеются ферменты, которые, действуя на мембраносвязанные субстраты, участвуют тем самым в биосинтезе мембран. С участием мембран в той или иной степени осуществляется большинство жизненно важных клеточных функций, например протекают такие разные процессы, как репликация прокариотической ДНК, биосинтез белков и их секреция, биоэнергетические процессы и функционирование систем гормонального ответа.
|
|
|
Данные, полученные при изучении клеток млекопитающих методом электронной микроскопии, свидетельствуют о наличии широко развитой сети внутриклеточных мембранных образований, которая занимает значительную часть внутреннего объема клетки. Сейчас уже не вызывает сомнений, что основные принципы структурной организации всех этих мембран по сути одинаковы. Более того, эти принципы соблюдаются также и в случае мембран растительных и бактериальных клеток. Основные закономерности, установленные Робертсоном в конце 1950-х гг., позволяют нам переносить результаты, полученные при исследовании одной мембранной системы, на другие системы. Естественно, учет специфики здесь необходим, поскольку, как это ни парадоксально звучит, одной из самых характерных особенностей мембран является их чрезвычайное разнообразие. Такое разнообразие обусловлено прежде всего разнообразием белков, присутствующих в каждой мембране, и способов их взаимодействия друг с другом и с компонентами цитоплазмы. Эти взаимодействия в конечном счете проявляются в специфической морфологии мембранных образований и могут быть связаны с латеральной гетерогенностью той или иной мембраны. Таким образом, основная задача заключается в том, чтобы, опираясь на общие представления о структуре и функциях мембран, выявить молекулярно-биологические основы их структурного и функционального разнообразия.
Успехов в исследовании мембран удалось достичь благодаря сравнительному изучению мембран из множества разнообразных организмов. Бактериальные клетки имеют довольно простую наружную оболочку, содержащую одну или две мембраны, которые можно модифицировать генетически или путем изменения условий роста клеток. Вирусы с оболочкой внедряются в клетки животных благодаря слиянию с плазматической мембраной последних и высвобождаются из клетки-хозяина, отпочковываясь от нее. Изучение созревания вирусных белков позволяет узнать много нового о процессах биосинтеза мембранных белков.
|
|
|
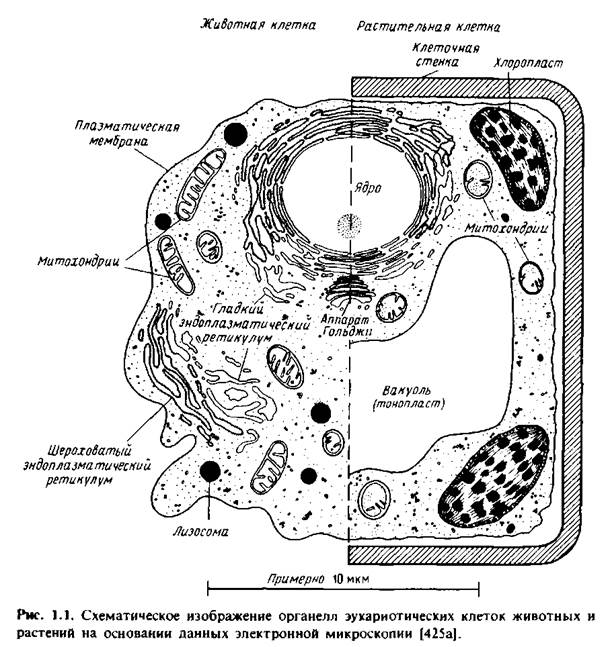
Эукариотические клетки содержат различные мембранные орга-неллы, причем каждая мембрана уникальна по своему составу, особенностям структурной организации и по характеру выполняемых функций. Для того чтобы понять мотивы исследований, описанных в последующих главах, необходимо получить некоторые общие представления о биологических функциях различных мембранных систем. На рис.1.1 схематически изображены мембраны, обычно представленные в животной и растительной клетках. Заметим, что внешний вид органелл неодинаков в клетках разного типа. Кроме того, некоторые клетки, например палочки сетчатки, а также клетки скелетных мышц, имеют высокоспециализированные мембраны, выполняющие уникальные функции.
Плазматическая мембрана. Плазматическая мембрана образует границу, на которой осуществляется контакт клетки с ее окружением.
Она содержит специализированные компоненты, участвующие в межклеточных контактах и взаимодействиях, в системах гормонального ответа и транспорта как малых, так и больших молекул из клетки и внутрь ее. Однако и сама плазматическая мембрана состоит из специализированных участков, которые имеют различное окружение. На рис.1.2 изображены апикальный и базолатеральный участки плазматической мембраны гепатоцитов и поляризованных эпителиальных клеток. Апикальная мембрана контактирует с какой-либо внутриклеточной средой. Так, у гепатоцитов она обращена в просвет желчных канальцев, а у эпителиальных клеток кишечника - в просвет желудочно-кишечного тракта
.
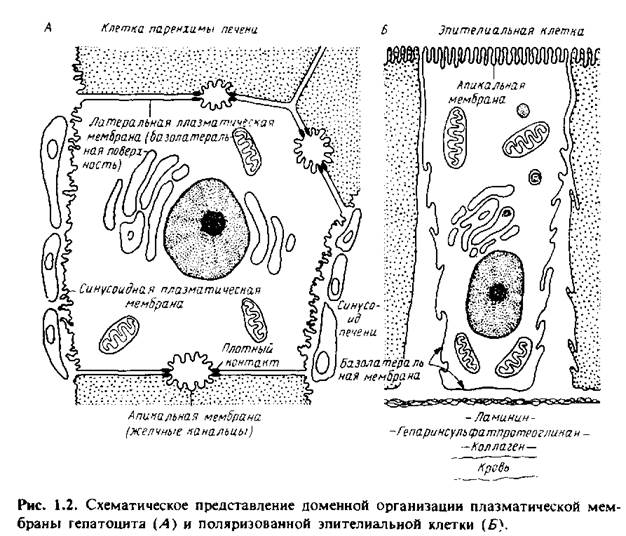
Она может иметь специализированные структуры, например микроворсинки; последние в некоторых всасывающих клетках образуют щеточную каемку. Микроворсинки значительно увеличивают площадь поверхности мембраны, в результате чего повышается эффективность мембранного транспорта. Базолатеральная мембрана находится в контакте с другими клетками или обращена в просвет кровеносных сосудов. Латеральная и синусоидная мембраны гепатоцитов различаются как по своей морфологии, так и биохимически.
Базолатеральная мембрана гепатоцитов имеет также специализированные структуры, ответственные за межклеточную адгезию и транспорт. Плотные контакты герметизируют область соприкосновения клеток и предотвращают перемешивание содержимого желчных канальцев и кровеносных сосудов.
|
|
|
Щелевые контакты содержат множество регулярно расположенных пор, которые позволяют небольшим молекулам проходить через плазматические мембраны двух соприкасающихся клеток. Электронно-микроскопические и биохимические исследования выявили характерные детали молекулярной организации этих пор, показав, что каждая из них содержит гексагонально упакованные белковые субъединицы.
Десмосомы также обеспечивают клеточную адгезию и участвуют во взаимодействии плазматической мембраны с элементами цитоскелета.
Апикальный, латеральный и синусоидный участки плазматической мембраны различаются морфологически и имеют уникальный состав и функции. Если клетки разрушить в мягких условиях, то можно выделить и очистить фракции, отвечающие этим участкам плазматической мембраны. Как на молекулярном уровне обеспечивается в клетке существование таких специализированных доменов, пока неясно, хотя известно, что не все их компоненты способны свободно диффундировать между доменами.
Ядерная мембрана. Ядерная оболочка клетки, находящейся в интерфазе, на электронных микрофотографиях имеет вид двух элементарных мембран с узким просветом между ними, называемым перинуклеарным пространством. Эта мембрана происходит из эндоплазматического ретикулума и, по-видимому, неразрывно связана с ним. Наиболее характерными морфологическими признаками являются порообразные структуры. Они имеют диаметр около 600 А и состоят из морфологически четко выявляемых компонентов, образующих октагональную решетку. В том месте, где расположены эти структуры, внутренняя и наружная ядерные мембраны выглядят слившимися. Полагают, что поры позволяют комплексам мРНК-белок переходить из ядра в цитоплазму, а регуляторным белкам перемещаться в обратном направлении, из цитоплазмы в ядро. Биохимические данные о ядерной оболочке весьма немногочисленны.
|
|
|
Эндоплазматический ретикулум. Это сложная сеть цистернообразных или трубчатых структур, которая занимает значительную часть внутреннего объема обычной животной клетки. Основная роль ЭР состоит в том, что он служит местом биосинтеза белков, которые затем секретируются, включаются в лизосомы или в плазматическую мембрану. Потенциально опасные для клетки гидролитические ферменты, которые должны секретироваться или накапливаться в лизосомах, подвергаются в ЭР процессингу до зрелой формы. С ЭР часто бывают связаны рибосомы, в результате чего на электронных микрофотографиях он выглядит шероховатым. Сложные процессы, в ходе которых осуществляется синтез белков, их превращение в зрелую форму и направленная доставка к месту назначения, описаны в гл.10.
Области ЭР, не содержащие рибосом, называются гладким ЭР. Здесь осуществляется биосинтез стеролов, протекают реакции детоксикации и происходит десатурация жирных кислот. Все эти процессы входят в сложную, согласованную систему транспорта электронов, осуществляемого при участии цитохромов bs и Р450.
Аппарат Гольджи. Эта органелла состоит из сети уплощенных мешков, собранных в стопки. Основная его функция заключается в посттрансляционной модификации гликопротеинов, синтезированных в эндоплазматическом ретикулуме и предназначенных для секреции, включения в плазматическую мембрану или доставки в лизосомы. Эти органеллы содержат гликозидазы и гликозилтрансферазы, которые вступают в действие последовательно, по мере того как белок, подвергаемый процессингу, перемещается от начала аппарата Гольджи до его конца. Фактически аппарат Гольджи состоит из совокупности отдельных мембран, образующих цистерны. Эти мембраны, которые можно выделить, характеризуются определенным набором ферментов. Механизмы транспорта мембран и секретируемых белков через аппарат Гольджи рассмотрены в гл.10.
Лизосомы. Эти органеллы ответственны за деградацию макромолекул и содержат ряд гидролитических ферментов, таких, как протеазы и липазы. Вещества, захваченные клеткой путем эндо- или фагоцитоза, которые необходимо расщепить, доставляются в лизосомы с помощью везикул. В лизосомах происходит также расщепление клеточных компонентов в ходе их обычного круговорота. Как осуществляются синтез лизосомных ферментов, их маркировка для доставки в лизосомы и последующий транспорт - изучено достаточно хорошо. Эти процессы рассматриваются в гл.10.
Пероксисомы. Эти органеллы содержат окислительные ферменты, участвующие в деградации малых молекул, таких, как аминокислоты, ксантин и, в особенности, жирные кислоты. Их название связано с присутствием в них каталазы, которая разлагает перекиси, образующиеся как побочные продукты при реакциях окисления.
Митохондрии. В этих органеллах осуществляется окислительное фосфорилирование, в результате чего в ходе окисления субстратов, таких, как NADH или сукцинат, образуется АТР. Митохондрии образованы двумя мембранами, разделенными некоторым промежутком. Внутренняя область митохондрий называется матриксом. Внутренняя мембрана образует складки в виде перегородок, называемых кристами, и содержит ферменты, участвующие в транспорте электронов и синтезе АТР. В гл.6 обсуждаются роль диффузии в плоскости мембраны компонентов цепи электронного транспорта и ее функциональное значение. Вопросы, связанные с синтезом белков митохондрий, который происходит в цитоплазме, и с их доставкой в один из митохондриальных компартментов или в одну из мембран, рассматриваются в гл.10.
Хлоропласты. Это органеллы, содержащие фотосинтетический аппарат. Они имеют наружную оболочку, образуемую двумя мембранами, и внутреннюю область - строму. В строме находятся тилакоидные мембраны, где локализованы компоненты системы фотосинтеза. На отдельных участках тилакоидные мембраны плотно упакованы в стопки, а на других обращены непосредственно к строме. Состав плотноупакованных и обращенных в строму доменов тилакоидной мембраны различен, что указывает на латеральную гетерогенность этой мембраны. Энзимология фотосинтезирующей цепи электронного транспорта обсуждается в разд.6.6.
Следует подчеркнуть, что при изучении каждой из мембран, упомянутых выше, а также и других специализированных мембран из клеток животных, растений или бактерий возникает целый комплекс важных и интересных вопросов, требующих своего решения, и открываются широкие возможности для биохимических исследований. Другие мембранные системы, представляющие интерес в этом отношении, будут описаны в последующих главах книги.
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 797; Нарушение авторских прав?; Мы поможем в написании вашей работы!