КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Гетерозисные скрещивания 1 страница
|
|
|
|
Трансгрессивные скрещивания
В результате трансгрессивных скрещиваний проявляется эффект суммирующего действия полимерных генов или так называемая трансгрессия, выражающаяся в устойчивом увеличении (положительная трансгрессия) или уменьшении (отрицательная трансгрессия) значения какого-либо полимерно наследующегося признака у отдельных гибридных особей F2 по сравнению с крайними значениями этого признака у родительских форм, т.е. перекрытие нормы проявления признаков.
Э. Ромедер и Г. Шенбах (1962) отмечают, что при селекции лесных древесных пород можно исходить из гипотезы, что ход роста контролируется многими аддитивно действующими генами. Для выяснения принципа можно принять, что продуктивность по массе обусловливается тремя одинаково действующими генами: W1-, W2-, W3- -, при этом их доминантные аллели W1, W2 и W3, обусловливают более высокую продуктивность, чем рецессивные w,, w2 и w3 Если в какой-либо популяции для скрещивания отбирают заметно выделяющиеся типы, то надеются, что они несут большее число доминантных генов продуктивности. Найти идеальный тип, обладающий всеми генами продуктивности в гомозиготном состоянии, для таких перекрестно опыляющихся растении, как лесные древесные, задача почти невозможная. Поэтому при выборе исходных форм для скрещиваний ценятся даже те особи, у которых хотя бы часть генов, обусловливающих продуктивность, находится в гомозиготном состоянии. Потомство таких скрещиваний будет расщепляться и показывать различную продуктивность. В результате, например, скрещивания двух генотипов W1 W1 W2 w2 W3 w3 и W1 w1 W2 W2 W3 W3 можно получить разное потомство, в том числе и с генотипом W1 W1 W2 W2 W3 W3, которое превосходит по продуктивности родительские деревья, т.е. обладает трансгрессией. Семенное потомство F2 этих отдельных особей в случае их самоопыления или переопыления между собой также отличается трансгрессией, поскольку оно гомозиготно по всем доминантным генам роста. Совокупность особей F2 отличается повышенным ростом как по сравнению с исходными родительскими формами, так и по сравнению с поколением F1. Объяснение явления трансгрессии близко к одной из гипотез гетерозиса — доминирования. Однако последний проявляется, как правило, лишь в поколении F1, а в следующих поколениях затухает.
|
|
|
Цель таких скрещиваний состоит в получении гибридного посадочного материала, обладающего гибридной мощностью, жизнеспособностью, повышенной продуктивностью или улучшенными качествами у гибридов первого поколения. Гетерозис считается истинным, если наблюдается превосходство гибрида по какому-нибудь признаку над признаком лучшего родителя, и гипотетическим, если превосходство наблюдается над средним значением признака обоих родителей.
А. Густафссон (1951, цит. по Р. Ригер и А. Михаэлис, 1967) под-, разделяет гетерозис по типу проявления на соматический, репродуктивный и приспособительный. Между этими тремя типами существуют переходы. Гетерозис соматический — это более мощное развитие вегетативных органов у гибридных растений; репродуктивный — более мощное развитие репродуктивных органов и повышенная фертильность, приводящие к формированию высокого урожая семян или плодов; приспособительный, или адаптивный, — повышение приспособленности гибридных организмов к изменяющимся условиям среды и их конкурентной способности в борьбе за существование.
Для объяснения причин гетерозиса разработано несколько гипотез. Наиболее распространена гипотеза доминирования, которая основана на представлении о том, что в процессе эволюции гены, благоприятно действующие на организм, становятся доминантными или полудоминантными, в то время как гены, действующие неблагоприятно, становятся рецессивными. Согласно этой гипотезе гетерозис объясняется тремя эффектами действия благоприятных доминантных генов: 1) подавление вредного действия рецессивных аллелей: Аа > аа; 2) аддитивный (суммирующий) эффект неаллель-ных доминантных генов, однонаправленно действующих на определенные количественные признаки, по которым в большинстве случаев и наблюдается гетерозис: А+В + С>А + В,А + Сили В + С >А, В или С (этот эффект присутствует и в случае трансгрессии, о чем было сказано выше); 3) комплементарное взаимодействие ряда не-аллельных доминантных генов: А <=> В > А + В.
|
|
|
Гипотеза сверхдоминирования объясняет гетерозис аллельным взаимодействием генов в гетерозиготном состоянии, вследствие чего АА < Аа > аа. Предполагается, что одинарная доза гена А благоприятнее действует на организм, чем его двойная доза в гомозиготе АА. Кроме того, действие разнонаправленных и независимых аллелей А и а при соединении их в гетерозиготу приобретает характер совместного однонаправленного доминирования; Ах а =* АА', АА < АА'> аа. Одним из доказательств обоснованности теории сверхдоминирования является факт моногибридного гетерозиса, проявляющегося при скрещивании двух гомозиготных линий, различия между которыми состоят только в генах одной аллельной пары: AABBCCDD х aaBBCCDD => AaBBCCDD. При этом АА... < Аа... > аа.... Гипотеза генетического баланса объясняет явление гетерозиса суммарным эффектом разнородных генетических процессов, изменяющих генетический баланс у гетерозиготы в сторону проявления той или иной формы гетерозиса.
Независимо от сущности разных теорий, объясняющих явление гетерозиса, с практической стороны важно иметь в виду, что гетерозис проявляется главным образом в Fv
Н.В. Старова (1980) у древесных пород выделяет три категории гетерозиса: популяционный, групповой и индивидуальный; а в каждой категории — типы гетерозиса: по характеру его проявления (генеративный, соматический, адаптивный) и по характеру взаимодействия генов (доминирование, сверхдоминирование, аддитивные и комплементарные эффекты). Поскольку описание типов было дано ранее, остановимся на характеристике категорий.
|
|
|
Популяционный гетерозис может возникать в результате длительной адаптивной эволюции в панмиктических естественных популяциях в результате удачных рекомбинаций и уравновешивающего естественного отбора, благоприятствующего гетерозиготам. Он может быть получен при географически отдаленной межпопуляционной гибридизации или при гибридизации различных природных изолятов в результате рекомбинации генов. В бывшем СССР были начаты работы по созданию плантаций сосны обыкновенной (И.Н. Патлай, П.И. Молотков), лиственницы (Ф.Д. Авров), березы (А.Я. Любавская) и других пород с использованием межпопуляционных скрещиваний географически отдаленных экотипов и изолятов.
Групповой гетерозис может быть получен при искусственной гибридизации родительских форм с высокой специфической комбинационной способностью, когда гибридная семья в целом по средним и максимальным показателям превосходит обе родительские формы. Примеры такого гетерозиса встречаются у тополей, ив и других пород.
Индивидуальный гетерозис наблюдается при межвидовых или географически отдаленных скрещиваниях, а также при скрещивании растений с различным уровнем плоидности. В этом случае не гибридные семьи в целом, а лишь отдельные экземпляры превосходят родительские формы (С.З. Курдиани). Индивидуальный гетерозис можно использовать в клоновой селекции при вегетативном и апомиктическом размножении гетерозисных форм для создания высокопродуктивных плантационных насаждений. Наиболее часто этот вид гетерозиса применяют при гибридизации тополей, ив, орехов рода Juglans L.
3.2. Методы гибридизации
Методы гибридизации зависят от биологических особенностей вида, характера исходного материала, требований к будущему сорту и др. Выделяют простые и сложные скрещивания, кроме того, существует ряд методов, сочетающих гибридизацию с другими подходами хозяйственного улучшения растений (инбридинг, полиплоидия, мутагенез и др.).
|
|
|
Простые скрещивания. Скрещивания между двумя родительскими формами, производимые однократно, называют простыми:
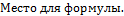
При простых скрещиваниях 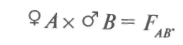 гибриды получаются от объединения наследственности двух родителей. Формообразовательный процесс в гибридных популяциях от простых скрещиваний идет на основе перераспределения наследственного материала, привнесенного в равном количестве одной парой родителей, поэтому простые скрещивания называют также парными. Разновидностью парных скрещиваний являются взаимные или реципрокные, когда в качестве материнского растения выступает растение, бывшее ранее отцовским:
гибриды получаются от объединения наследственности двух родителей. Формообразовательный процесс в гибридных популяциях от простых скрещиваний идет на основе перераспределения наследственного материала, привнесенного в равном количестве одной парой родителей, поэтому простые скрещивания называют также парными. Разновидностью парных скрещиваний являются взаимные или реципрокные, когда в качестве материнского растения выступает растение, бывшее ранее отцовским:
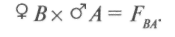
Реципрокные скрещивания применяются:
а) когда наследование какого-либо важного хозяйственно-биологического признака (например, зимостойкости, роста и др.) связано с цитоплазмой; гибриды наследуют это свойство сильнее в том случае, если носитель ценного признака берется в виде материнского растения;
б) когда завязываемость семян зависит от того, в качестве материнской или отцовской берется та или иная форма.
Сложные скрещивания. В сложных скрещиваниях участвуют более двух родительских форм, или гибридное потомство повторно скрещивается с одним из родителей. Они делятся на ступенчатые и возвратные. Ступенчатые скрещивания применяются, когда необходимо соединить в гибридном потомстве наследственность нескольких родительских форм. Их можно представить в виде следующих простейших формул:
1)[(АхВ)хС]х D; 2)[(AxB)x(Cx D)] х Е.
В первом случае гибрид, полученный от скрещивания двух родительских форм А и В, дополнительно скрещивается с формой С, а затем с формой D; в целом здесь объединяется наследственность четырех родительских форм. Во втором случае сначала скрещиваются попарно формы А и В, С и D, а их гибридное потомство скрещивается между собой и с формой Е; в целом здесь объединяется наследственность пяти родительских форм. В обоих случаях скрещивания осуществляются последовательно, ступенчато.
Возвратные скрещивания — это такие скрещивания, при которых гибрид повторно скрещивается с одним из родителей. Их применяют в двух случаях: 1) для преодоления бесплодия гибридов первого поколения при отдаленной гибридизации: (Ах В) х В; 2) для усиления в гибридном потомстве желаемых свойств одного из родителей. В этом случае возвратные скрещивания называют насыщающими. Чаще всего их используют при выведении сортов, устойчивых к болезням или неблагоприятным абиотическим факторам среды. Лучший высокопродуктивный, но неустойчивый к действию какого - либо неблагоприятного фактора сорт берется в качестве отцовской формы, а сорт, обладающий устойчивостью к нему, — в качестве материнской. Например, скрещиваются устойчивый сорт А с быстрорастущим, но мало устойчивым сортом В, тогда мы можем иметь следующую последовательность возвратных скрещиваний, каждое из которых называется беккроссом (табл. 3.2). В результате первого беккросса количество отцовского ядерного материала увеличивается до 75%, после пятого — оно равняется 99,2%, т.е. происходит почти полное поглощение материнской наследственности отцовской, поэтому такие насыщающие скрещивания называются поглотительными.
Использование в гибридизации явления инбридинга позволило разработать инцухт-гетерозисный метод получения гетерозисных растений. При этом методе сначала получают инцухт-линии или инбред -линии (самоопыленные линии), представляющие собой потомство одного перекрестно опыляющегося растения, полученное в результате принудительного самоопыления. Исходное растение, которое принудительно подвергли самоопылению, обозначается символом 1о, первое его инбредное потомство — символом 11, второе — символом 1 2 и т.д. Затем отбирают инбредные линии, обладающие высокой комбинационной способностью, и скрещивают их между собой. В результате получают гибриды, обладающие повышенным гетерозисом. Такие гибриды называют инцухт-гетерозисными. Прекрасные результаты применения этого метода получены на сельскохозяйственных растениях, в частности у кукурузы. О его применимости для лесных пород указывал еще А.И. Колесников, а в более позднее время — Э. Ромедер и Г. Шенбах (1962).
Таблица 3.2
Схема применения насыщающих скрещиваний в селекции на устойчивость (по Г.В. Гуляеву, А.П. Дубинину, 1974)
| Номер беккросса | Последовательность скрещиваний | Процент насыщения устойчивого сорта свойствами лучшего высокопродуктивного сорта |
| А х В = F1; | ||
| F1*B = F2 | ||
| F2*B = F3 | 87,5 | |
| F3*B=F4 | 93,7 | |
| F4 *B = F5 | 98,4 | |
| F5*B = F6 | 99,2 |
Необходимо отметить, что в зависимости от способа размножения те или иные методы гибридизации и селекции дают более или менее важные практические результаты.
Так, для самоопылителей наибольшее распространение получили методы комбинационной селекции, как простые, так и сложные, особенно метод ступенчатой и отдаленной гибридизации. Среди перспективных методов можно отметить: использование принципов генетически регулируемого гетерозиса; получение индуцированных мутаций при применении радиации и химических мутагенов и использование искусственно получаемых аллополиплоидов (Н.П. Дубинин, Я.Л. Глембоцкий, 1967).
В селекции перекрестников сельскохозяйственных культур исторически прослеживается переход от массового отбора к семейственному отбору и затем к получению самоопыленных линий и использованию гетерозисных гибридов. Здесь, как и при скрещивании самоопылителей, наблюдаются простые и сложные скрещивания. Новыми методами в этом случае являются скрещивание инцухт-ли-ний (межлинейные гибриды) или скрещивания инцухт-линий на сорт (сортолинейные гибриды), при которых проявляется гетерозис.
В селекции вегетативно размножаемых растений наибольшее значение приобрели такие методы селекции, как отдаленная гибридизация, экспериментальное получение полиплоидов и искусственный мутагенез. При всех этих методах важно получить хотя бы одно растение с улучшенными хозяйственными признаками, которое в последующем можно размножить вегетативным путем. Успех здесь зависит в основном от выбора исходного материала и технического осуществления методов.
3.3. Техника гибридизации
Проведение контролируемых скрещиваний лесных деревьев — очень дорогое и долговременное мероприятие. Поэтому прежде чем его проводить, составляют схему (план) скрещиваний, при разработке которой стараются сделать так, чтобы можно было одновременно достичь нескольких целей. Различают самоопыление, системы скрещиваний с неизвестными отцами (свободное опыление и поликросс) и системы скрещивания с известными отцами. Планы скрещиваний составляют, собственно говоря, только для последней группы скрещиваний. Различают полную схему диаллельных скрещиваний, модифицированную схему диаллельных скрещиваний, мистичные схемы диаллельных скрещиваний, факториальную схему скрещиваний, простые скрещивания (В. Zobel, J. Talbert, 1984, И. Ditlevsen, 1985).
Полная схема диаллельных скрещиваний (рис. 3.1) считается одной из лучших, поскольку включает все возможные варианты скрещивания и дает почти полную информацию о генетических характеристиках изучаемых клонов. Схема может давать информацию об ОКС и СКС и их дисперсиях. Материал также создает лучшую стартовую точку для отбора наилучших индивидуумов или пар клонов, подходящих для создания биклоновых плантаций. Схема, к сожалению, очень трудна для осуществления, особенно с экономической точки зрения. Например, для реализации полной диаллельной схемы для 20 клонов надо провести 400 контролируемых скрещиваний или 380, если исключить самоопыление. Поэтому в практике используют другие схемы (рис. 3.2-3.4).
Модифицированная схема диаллельных скрещиваний представляет собой ограниченную диаллельную схему скрещиваний. В этом случае из полной схемы исключаются реципрокные скрещивания и самоопыления (рис. 3.2), что значительно удешевляет реализацию схемы. Эта схема дает примерно такую же информацию, как и полная схема диаллельных скрещиваний, но ограничения, допущенные в схеме, не гарантируют такой же точности в опытах при определении параметров.
Частичная схема диаллельных скрещиваний может отличаться от полной и модифицированной схемы диаллельных скрещиваний настолько, что один клон не скрещивается со всеми другими клонами (рис. 3.3). Схема менее эффективна, чем схемы полных и модифицированных диаллельных скрещиваний. Однако это компенсируется тем, что большое количество потомства может быть испытано при довольно низких затратах. К сожалению, отсутствие оценок, которые нельзя получить в частичных схемах, значительно усложняют расчеты.
| Матери | Отцы | |||||||||
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X | |
| X | X | X | X | X | X | X | X | X | X |
Рис. 3.1. Полная схема диаллельных скрещиваний: X — самоопыление
| Матери | Отцы | ||||||||
| X | |||||||||
| X | X | ||||||||
| X | X | X | |||||||
| X | X | X | X | ||||||
| X | X | X | X | X | |||||
| X | X | X | X | X | X | ||||
| X | X | X | X | X | X | X | |||
| X | X | X | X | X | X | X | X | ||
| X | X | X | X | X | X | X | X | X |
Рис. 3.2. Модифицированная схема диаллельных скрещиваний
| Матери | Отцы | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X | ||||||||||
| X | X |
Рис. 3.3. Частичная диаллельная схема Брауна
Факториальная схема скрещивания отличается тем, что в ней все материнские клоны скрещиваются с одними и теми же отцовскими клонами (рис. 3.4). Часто это может быть небольшое число отцовских растений, называемых также общими тестерами. Данная схема может также рассматриваться как разновидность полной диаллельной схемы, включающей все комбинации одной группы матерей и другой группы отцов. Схема очень распространена в США под названием Северо-Каролинская II. Нормально четыре различных отца используются для схемы скрещивания, но, как правило, число отцов в схеме зависит от значения специфических комбинационных эффектов. Так как схема часто включает очень немногое количество отцов и одни и те же клоны не являются одновременно материнскими и отцовскими, то трудно сравнить ОКС каждого родителя. Трудно также отобрать потомство для следующего поколения семенных плантаций, особенно если используется только несколько отцов, являющихся часто уже родственными друг другу. Преимущество этой схемы в простоте ее выполнения и относительной дешевизне; в то же время анализ результатов легче.
| Матери | Отцы | |||
| X | X | X | X | |
| X | X | X | X " | |
| X | X | X | X | |
| X | X | X | X | |
| X | X | X | X | |
| X | X | X | X | |
| X | X | X | X | |
| X | X | X | X |
Рис. 3.4. Факториальная схема скрещивания
Простое скрещивание пар характеризуется тем, что в этом случае каждый клон включен только один раз — как мать или как отец. Эта схема особенно хороша, если целью является создание популяции для отбора особей для новых семенных плантаций или для использования в дальнейшей селекционной работе. Другим преимуществом использования простого скрещивания пар является то, что большое количество клонов может быть испытано при одинаковой схеме, и обычно очень дешево может быть получено потомство на базе контролируемых скрещиваний. С другой стороны, возможности оценки ОКС и дисперсии ОКС и СКС обычно не очень хороши. Выбор соответствующей схемы зависит от цели контролируемых скрещивании. В табл. 3.3 суммированы достоинства и недостатки названных схем.
При планировании скрещиваний следует учитывать ряд особенностей исходного материала, отмеченных С.С. Пятницким (1961):
• для получения крепкого потомства необходимо отбирать для скрещивания здоровые неистощенные экземпляры;
Таблица 3.3
Сравнение схем скрещиваний
(по В. Ditlevsen, 1985)
| Система скрещивания | Оценка ОКС | Отбор плюсовых деревьев | Затраты | Оценка дисперсии ОКС и СКС |
| Свободное опыление | Неплохая | Возможен, но не эффективен | Низкие | Затруднена |
| Поликросс | Очень хорошая | Очень незначителен и только, если дисперсия близкородственного скрещивания ослаблена | Очень низкие | Хорошая оценка дисперсии ОКС |
| Полная диаллельная | Превосходная | Превосходный | Очень высокие | Превосходная |
| Модифицированная диаллельная | Превосходная | Превосходный | Очень высокие | Очень хорошая |
| Частичная диаллельная | Хорошая | Очень хороший | Достаточные | Хорошая оценка дисперсии ОКС. Дисперсия СКС может быть определена, но это трудно с точки зрения обработки данных |
| Факториальная | Хорошая | Только в немногих случаях и тогда, когда дисперсия близкородственного скрещивания низкая | Достаточные | Хорошая |
| Простое скрещи-h.niHc пар | Плохая | Хороший | Очень низкие | Плохая |
• у молодых растений выход гибридных семян и плодов выше, чем у старых;
• при скрещивании с местными видами полученные гибриды больше уклоняются в их сторону;
• при скрещивании культурной формы с дикой уклонение идет в сторону последней;
• при скрещиваниях материалов из географически отдаленных местностей или экологически неоднородных условий местопроизрастания больше вероятности получить хозяйственно ценные гибриды;
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 1255; Нарушение авторских прав?; Мы поможем в написании вашей работы!