КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Часть III. Историческое развитие (филогенез) организмов 9 страница
|
|
|
|
Австралия уже со времени распада Гондваны занимала обособленное положение, будучи связана с другими материками, главным образом, через Антарктиду (см. рис. 60). Поэтому в Австралию ко времени ее изоляции, вероятно, еще не проникли плацентарные млекопитающие. В Австралии сформировалась уникальная фауна сумчатых и однопроходных, отчасти сохранившаяся до наших дней. Однако палентологическая история австралийских млекопитающих известна еще очень плохо. Вероятно, современная фауна этого материка значительно обеднена по сравнению с предшествовавшими эпохами кайнозоя. Относительно хорошо известна поздняя - плейстоценовая - фауна Австралии. Среди плейстоценовых сумчатых здесь можно упомянуть дипротодона (Diprotodon) - самого крупного среди известных сумчатых, достигавшего размеров носорога с черепом около 1 м и мощными бивнеподобными резцами в верхней и нижней челюстях. Очень своеобразен был "сумчатый лев" (Thylacoleo), у которого коренные зубы были редуцированы, зато имелись огромные режущие предкоренные. Некоторые палеонтологи считают
;это животное хищником, другие же полагают, что "сумчатый лев" питался какими-то плодами.
В Южную Америку до ее отделения от Северной Америки успели проникнуть некоторые архаические группы плацентарных млекопитающих, имелись там и сумчатые. На основе этих групп в течение длительной изоляции Южной Америки сложилась удивительная фауна архаических млекопитающих. В Северной Америке, Европе и Азии в течение палеогена и неогена плацентарные млекопитающие в целом сделали большой шаг вперед по пути морфофизиологического прогресса (увеличение и усовершенствование головного мозга, усложнение поведения, прогрессивные изменения локомоторного аппарата, органов пищеварения, зубной системы и т. д.). В Южной Америке все это время продолжали процветать архаические группы, сохранявшие примитивное строение головного мозга, но также на свой лад совершенствовавшие органы пищеварения и локомоции, причем многие их адаптации оказались очень сходными с соответствующими приспособления-Ми млекопитающих Старого Света и Северной Америки.
Очень разнообразны и причудливы были южноамериканские растительноядные млекопитающие. Копытные были представлены группами нотоунгулят (Notoungulata), астрапотериев (Astra-Potheria), литоптерн (Litopterna) и пиротериев (Pyrotheria). Нотоунгуляты известны также из палеоцена Азии (где, вероятно, и возникла эта группа) и раннего эоцена Северной Америки, но исчезли повсеместно уже в эоцене, сохранившись и процветая лишь в Южной Америке. Южноамериканские нотоунгуляты были очень разнообразны; среди них были и проворные мелкие животные, подобные типотериям (Typotheria), с длинными задними конечностями, которые напоминали кроликов пропорциями тела, размерами и строением зубов; и тяжелые коротконогие токсо-донты (Toxodonta), достигавшие 2,75 м длины и похожие на некоторых безрогих носорогов.
Астрапотерии и пиротерии были крупнейшими копытными Южной Америки, достигая размеров крупных носорогов и даже слонов. С последними они были сходны также строением конечностей и наличием хобота, а пиротерии удивительно напоминали настоящих хоботных еще и обликом черепа и зубной системы (вплоть до крупных бивнеподобных резцов в верхней и нижней челюстях).
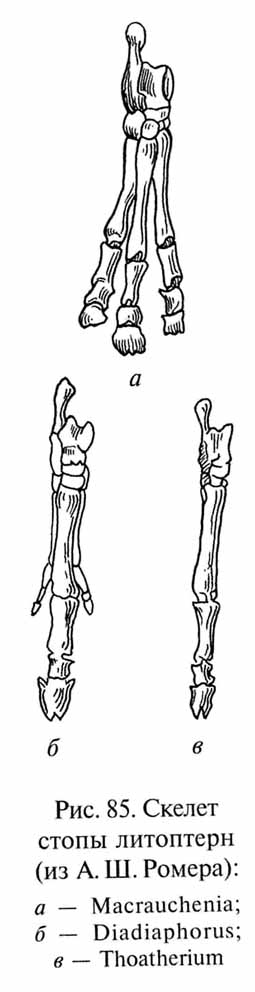
В эволюции литоптерн возникли формы, конечности которых были поразительно сходны с лошадиными (яркий пример эволюционного параллелизма). В процессе приспособления к быстрому бегу по относительно плотной почве саванн у этих литоптерн усиливался средний палец и редуцировались боковые (рис. 85), вплоть до полной их редукции у миоценового тоатерия (Thoatherium). Тоатерий был в этом отношении даже более специализирован, чем настоящие лошади, сохраняющие рудименты боковых пальцев в виде так называемых грифельных косточек (см. рис. 81). Наиболее поздний представитель литоптерн - плейстоценовая макраухения (Macrauchenia) размерами и пропорциями тела напоминала верблюда, но имела трехпалые ноги, а ноздри на черепе располагались сверху, как у хоботных животных (возможно, макраухения также имела небольшой хобот).
|
|
|
|
|
|
Таким образом, при отсутствии в Южной Америке настоящих хоботных, носорогов, лошадей, зайцеобразных и других групп млекопитающих, эволюционировавших в Евразии и Северной Америке, там конвергентно возникли формы, удивительно напоминавшие внешне своих аналогов.
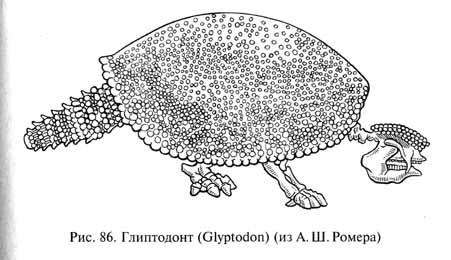
Зато совершенно специфичны южноамериканские неполнозубые (Edentata). Эта группа, возможно, возникла в Северной Америке, откуда известны остатки древнейших палеоценовых неполнозубых, но центром ее эволюции стала Южная Америка в условиях географической изоляции. И современные неполнозубые (ленивцы, броненосцы, муравьеды) обитают в основном в Южной Америке, за исключением нескольких видов, проникших в южную часть Северной Америки после восстановления ее связи с Южной. Среди вымерших неполнозубых интересны родственные броненосцам глиптодонты (Glyptodontidae, рис. 86), достигавшие до 5 м в длину. Их тело было одето сплошным панцирем из сросшихся мелких костных пластинок - остеодерм. "Шапочка" из остеодерм прикрывала сверху голову. Пальцы коротких мощных лап были защищены копытами.

Но особенно интересной группой неполнозубых были гигантские наземные ленивцы, или мегатерии (Megatheriidae, рис. 87). Эти животные, как и глиптодонты, известны начиная с олигоце-на. Некоторые более поздние представители наземных ленивцев достигали огромных размеров: плейстоценовый мегатерий (Megatherium) был высотой со слона при длине тела около 6 м. Внешний облик этих странных существ был необычен. Огромное тело опиралось на мощные конечности, особенно велики были задние лапы с большими широкими ступнями. Передние конечности несли крупные изогнутые когти и при ходьбе опирались не на ладонную, а на боковой край тыльной стороны, сгибая пальцы внутрь. Вероятно, эти гиганты передвигались медленно и довольно неуклюже. Судя по строению их скелета, мегатерии могли вздымать свое огромное тело вертикально, опираясь на задние лапы; передними конечностями они при этом пригибали ветви и кроны деревьев вниз, цепляясь крепкими когтями. Питались гигантские ленивцы листьями. Конечно, не все мегатерии были столь велики, меньшие виды имели длину тела примерно 2-3 м. Гигантские ленивцы были широко распространены в Южной Америке, а после соединения двух Америк проникли и в Северную.
Плацентарных хищников в Южной Америке не было. Соответствующие экологические ниши занимали в основном разнообразные сумчатые хищники (Borhyaenidae), коротконогие и большеголовые звери, вооруженные острыми клыками и крепкими когтями; самые крупные виды достигали размеров медведей. С сумчатыми хищниками конкурировали упомянутые нами выше полутора - трехметровые хищные нелетающие птицы - фороракосы, а также самые крупные из известных летающих птиц - тераторнисы (Teratornithidae). Среди них аргентавус (Argentavus), известный из верхнемиоценовых отложений Аргентины, достигал в размахе крыльев 7,5 м при вероятной массе тела около 80 кг. Череп аргентавуса (длиной до 55 см) и других тераторнисов был вооружен мощным клювом. Вероятно, эти птицы могли проглатывать целиком добычу размером с зайца.
Существует предположение, что в Южной Америке вернулись к наземной жизни и крокодилы из группы себекозухий (Sebecosuchia), которые стали новой группой сухопутных хищных архозавров. У себекозухий череп был сжат с боков, а не уплощен сверху, как у большинства крокодилов, плоский череп которых с обращенными вверх глазницами и ноздрями характерен для хищников, подстерегающих в воде наземную добычу.
В плиоценовую эпоху неогена вновь восстановилась связь между Южной и Северной Америками, и по сухопутному "мосту" в Южную Америку хлынула волна новых, более современных групп млекопитающих (непарно- и парнокопытные, зайцеобразные, хоботные, хищники-фиссипедии). Это привело к быстрому вымиранию большинства специфических южноамериканских групп растительноядных и хищных млекопитающих. До плейстоцена продержались лишь немногие литоптерны и нотоунгуляты. Зато неполнозубые оказались вполне конкурентоспособными по отношению к высшим группам млекопитающих и даже расселились в Северную Америку. Это относится и к глиптодонтам, и к гигантским ленивцам, обитавшим в плейстоцене на огромных территориях от Аризоны и Флориды до Аргентины. Вселились в Северную Америку и гигантские хищные птицы - тераторнисы. Гигантские ленивцы дожили, несомненно, до времени появления в Америке человека. В некоторых пещерах в Северной и Южной Америке сохранились не только скелеты этих животных, но и остатки их шкур - вместе с каменными и костяными орудиями человека. Возможно, последние мегатерии вымерли всего несколько тысячелетий назад. Не исключено, что преследование со стороны людей послужило одной из причин вымирания этих медлительных и неуклюжих животных. В то же время, забегая несколько вперед, отметим, что в конце плейстоцена в обеих Америках вымерло подавляющее большинство существовавших здесь прежде многочисленных крупных млекопитающих.
Развитие жизни в неогене
Как мы уже упоминали, в течение неогена произошло значительное повышение материков, сопровождавшееся морской регрессией, постепенным иссушением климата и прогрессирующим развитием открытых ландшафтов. Одновременно происходило постепенное понижение средних температур на поверхности Земли. Эти изменения природных условий оказали глубокое влияние на флору (см. начало этой главы) и фауну.
В неогене получили преобладание виды наземных животных, приспособленные к жизни в открытых и относительно сухих местообитаниях типа лесостепей, степей и саванн. В олигоцене вымерли многие виды, приспособленные к жизни в лесах, на влажных лугах, заболоченных участках и других подобных биотопах, характерных для палеогена. Эта судьба постигла многие группы непарнокопытных. Зато парнокопытные, начиная с олигоцена, широко распространились и быстро прогрессировали. Особенно это относится к наиболее новым их группам: оленям (Cervidae), жирафам (Giraffidae), вилорогам (Antilocapridae) и многочисленным полорогим (Bovidae). Успех этих групп прежде всего связан со специализацией зубов и пищеварительной системы к переработке грубых растительных кормов - злаков и других растений степей и саванн, а также с приспособлениями к быстрому бегу по относительно плотной почве травянистых равнин.
Из непарнокопытных в неогене успешно конкурировали с парнокопытными лишь лошади и носороги. Миоценовые и плиоценовые лошади были очень многочисленны и разнообразны. Обшее направление их эволюции (как и обычно, прослеживающееся в нескольких независимых и параллельных филетических линиях) характеризовалось дальнейшим увеличением размеров тела, происходившим коррелятивно удлинением морды и соответственно зубного ряда, увеличением размеров коренных зубов и развитием сложной системы гребней на их жующей поверхности, а также удлинением конечностей с дальнейшей редукцией боковых пальцев (см. рис. 81). При этом в разных филетических линиях преобразования различных указанных признаков происходили с разной скоростью и независимо друг от друга: в одних линиях быстрее прогрессировали преобразования зубов, в других - конечностей и т.п.
На основании анализа этих преобразований Г.Осборном был сформулирован так называемый принцип независимой эволюции различных признаков, согласно которому разные признаки организма могут изменяться в ходе эволюции в определенной степени независимо друг от друга. Разумеется, независимо эволюционировать могут лишь те признаки, которые не связаны друг с другом какими-либо коррелятивными взаимоотношениями. Принцип Осборна лежит в основе концепции "мозаичной эволюции" Г. де Вира (см. с. 249, 350).
Для миоцена были характерны трехпалые лошади (Parahippus, Merychippus, Hipparion и др.), у которых боковые пальцы были укорочены и касались субстрата, вероятно, лишь при более медленной ходьбе и при пересечении участков с влажной почвой, препятствуя увязанию. Центром эволюции лошадей оставалась Северная Америка, откуда эти непарнокопытные проникли в Евразию. В плиоцене появились формы (Pliohippus, Plesippus), строение конечностей и зубов у которых приближалось к таковым у современной лошади (Equus), известной начиная с плейстоцена. Отметим, что однопалость развивалась независимо и параллельно по меньшей мере в четырех обособленных филетических линиях лошадей (Astrohippus, Pliohippus, Hippidion - Onohippidion, Protohippus - Equus).
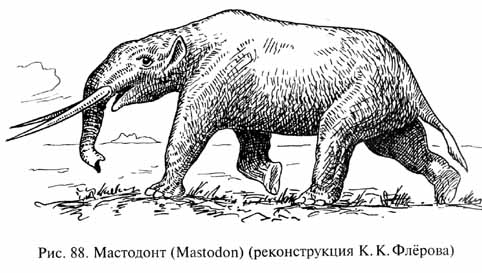
В неогене максимального расцвета достигли хоботные, расселившиеся из Африки в Евразию, затем в Северную и, наконец, в Южную Америку. Центральный эволюционный ствол хоботных был представлен мастодонтами (рис. 88), среди которых появились очень разнообразные формы: с коротким и длинным хоботом, различного строения бивнями и т.п. Причудливо выглядели лопаторылые мастодонты (Amebelodon, Platybelodon), у которых передняя часть нижней челюсти чрезвычайно удлинилась, образуя вместе с огромными уплощенными резцами своего рода "совок" или "лопату", выдвинутую вперед и достигавшую чудовищных размеров: животное высотой в холке 2,5 м имело нижнюю челюсть длиной более 2 м. По-видимому, лопаторылые мастодонты использовали свою необычайную нижнюю челюсть именно как лопату, подцепляя ею корм - может быть, со дна неглубоких полузаболоченных водоемов (сочные листья, клубни или корневодных растений).
|
|
|
|
|
|
Боковой ветвью эволюционного ствола мастодонтов были миоценовые динотерии (Deinotherium), у которых передняя часть нижней челюсти была отогнута вниз, а сидящие на ней бивни были направлены вниз и назад. Сейчас трудно представить, как использовали такие бивни эти крупные хоботные.
В миоцене от мастодонтов возникли настоящие слоны (Elephantidae). Их родиной, вероятно, была Южная Азия, откуда они в плиоцене расселились в Европу, Северную Америку и Африку. У слонов в отличие от мастодонтов и динотериев сохранилась лишь верхнечелюстная пара бивней, но особенно характерной чертой настоящих слонов является строение огромных коренных зубов. Их высокие и сжатые с боков коронки состоят как бы из многочисленных поперечных пластин, образующих на жевательной поверхности узор из узких поперечных гребней и желобков.
Обилию растительноядных млекопитающих в лесостепях и саваннах неогена соответствует и разнообразие хищников. В миоцене существовали уже все семейства современных хищных млекопитающих, большинство современных родов известно начиная с плейстоцена.

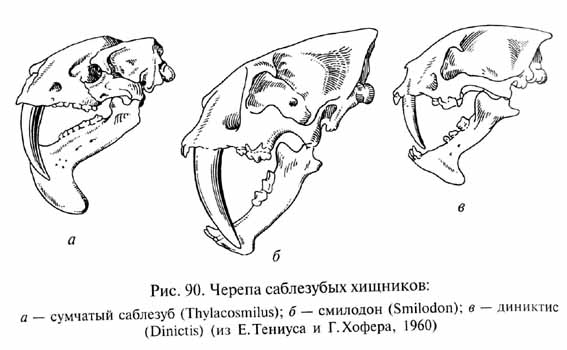
Среди ископаемых кошачьих (Felidae) интересна группа так называемых саблезубов (или правильнее "кинжалозубов" - Маchairodontinae), существовавшая с олигоцена до плейстоцена. Этих хищников иногда называют "саблезубыми тиграми", но к тиграм они были не более близки, чем к львам, леопардам или домашним кошкам (которые все являются представителями другого подсемейства кошачьих, собственно кошек - Felinae). Саблезубы (рис. 89) обладали огромными уплощенными с боков клыками, достигавшими у крупных форм длины 15 см; этими клыкам хищники и обязаны своим названием. Это были крепкие коренастые животные с мощной шеей и могучими когтистыми лапами, причем передние конечности были длиннее и сильнее задних. Такой тип телосложения совсем не соответствует способам охоты, характерным для современных кошачьих: едва ли саблезубы могли совершать дальние прыжки на добычу из засады или стремительно преследовать ее даже на коротких расстояниях. Огромные клыки и способность чрезвычайно широко открывать пасть, отводя нижнюю челюсть вниз и назад под углом, близким к прямому или даже несколько большим, указывают на то, что главным способом умерщвления добычи у саблезубов было нанесение сильных ударов клыками, глубоко вонзавшимися в тело жертвы и причинявшими тяжелые ранения. Вероятно, основной добычей крупных саблезубов (отметим, что среди них имелись и относительно мелкие виды) были крупные толстокожие растительноядные животные (хоботные, носороги и т.п.), на которых хищники нападали из засады.
Интересно, что саблезубые формы хищников возникали неоднократно и в разное время среди различных групп млекопитающих. Древнейшими из них являются эоценовые саблезубые креодонты Machaeroides - относительно мелкие хищники, питавшиеся, вероятно, кондиляртрами. В плиоцене в Южной Америке существовал сумчатый саблезуб (Thylacosmilus, рис. 90). Его добычей, вероятно, были такие крупные толстокожие животные, как токсодонты, мегатерии и т.п. В плейстоцене его вытеснили настоящие саблезубы (Smilodon), проникшие в Южную Америку из Северной вместе с другими видами высших млекопитающих. Возникновение саблезубых хищников в разных группах млекопитающих является одним из ярких примеров параллельной эволюции независимых филетических линий. Вымирание большинства крупных растительноядных форм к концу плейстоцена привело к вымиранию охотившихся на них саблезубов.
В неогене водные млекопитающие пополнились ластоногими (Pinnipedia), ископаемые остатки древнейшего представителя которых (Enaliarctos) известны из пограничных отложений олиго-цена и миоцена. Возможно, разные филогенетические стволы ластоногих возникли от примитивных представителей различных семейств наземных хищников: ушастые тюлени (Otariidae) и моржи (Odobaenidae) - от медвежьих, а настоящие или безухие тюлени (Phocidae) - от куньих. Общие особенности строения ластоногих сформировались параллельно и независимо в этих двух филогенетических стволах.
На огромных пространствах Северной Америки, Евразии и Африки, занятых лесостепями и степями с разной степенью увлажненности, в позднем миоцене и плиоцене получила широкое распространение фауна, которую называют "гиппарионовой" - по самому характерному ее представителю, трехпалой лошади - гиппариону (Hipparion, рис. 91). Многотысячные табуны гиппарионов паслись на лугах и остепненных участках, перемежающихся с рощами широколиственных листопадных деревьев и кустарников (дуб, бук, ясень, вяз, ольха, различные виды тополя, граб, клен, грецкий орех и др.) и разреженными лесами, включавшими и широколиственные, и хвойные деревья (сосны, секвойи, болотные кипарисы), прорезанных долинами рек и заболоченными низинами. Вместе с гиппарионами обитали разнообразные антилопы, олени, древние жирафы (например, палеотрагусы Palaeotragus, похожие на современных окапи, и тяжелые буйволообразные сиватерии - Sivatherium), хоботные (мастодонты и динотерии), носороги и страусы, близкие к современным африканским. В состав гиппарионовой фауны входили, конечно, и хищники (саблезубы, различные псовые, гиены, росомахи, куницы, виверры и др.), а также грызуны, зайцеобразные и обезьяны1.
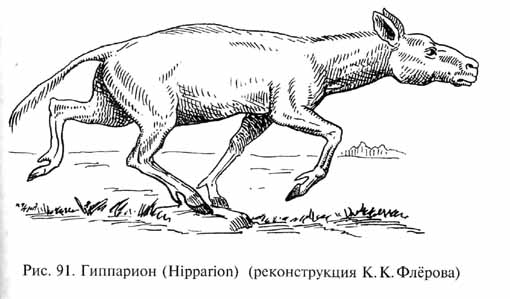
Гиппарионовая фауна, рассматриваемая в целом, была в значительной степени предковой для современной фауны Африки, включающей представителей тех же основных групп животных. По аналогии с недавним прошлым африканских саванн и по обилию ископаемых остатков различных копытных в местонахождениях гиппарионовой фауны можно заключить, что в конце миоцена - плиоцене на огромных пространствах равнин Старого и Нового Света паслись неисчислимые стада гиппарионов, антилоп, оленей, хоботных и других растительноядных млекопитающих. Ю.А.Орлов высказал предположение, что в зоне умеренного пояса, где уже в неогене были отчетливо выражены сезонные изменения климата, эти гигантские стада копытных и сопровождающие их хищники должны были совершать весной и осенью далекие миграции, подобные миграциям африканских копытных, но в гораздо большем масштабе.
Четвертичный период
В плейстоцене происходило дальнейшее похолодание климата, сопровождавшееся постепенным отступанием теплолюбивой флоры и фауны в Евразии и Северной Америке к югу. Широкое распространение получили листопадные и хвойные леса, общий облик которых уже напоминал современные леса умеренного пояса. Состав фауны также приближался к современному; появились многие современные роды млекопитающих.
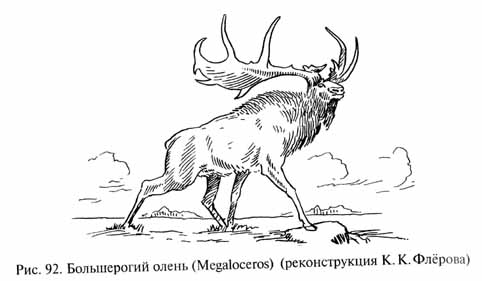
Однако в плейстоцене еще существовал целый ряд более древних форм и групп животных (мегатерии, глиптодонты, макраухении, токсодонты, мастодонты, динотерии, саблезубы и др.), особенно многочисленные в Америке. Появились и некоторые своеобразные виды, которым не было суждено пережить плейстоценовую эпоху. Среди них интересны гигантские бобры - трогон-терии (Trogontherium), имевшие более 2 м длины; носороги - эласмотерии (Elasmotherium), обладавшие огромным вздутием лобной области черепа, адаптивная роль которого остается неизвестной; большерогие олени - мегалоцеросы, или "ирландские лоси" (Megaloceros; рис. 92), достигавшие высоты в холке около 2 м, но особенно замечательные своими гигантскими ветвистыми рогами, размах которых превышал 4 м. Эти поразительные рога требовали, конечно, от организма немалых энергетических затрат и на их развитие в онтогенезе, и на поддержание головы с таким тяжелым вооружением. Вероятно, мегалоцеросы использовали свои рога для тех же целей, что и все остальные олени: как демонстрационный признак и "турнирное оружие" в борьбе между самцами в период гона (у самок рога отсутствовали). Развитие рогов было обусловлено половым отбором, который нередко (при отсутствии опасных хищников или при хорошей защищенности данного вида особенностями его образа жизни занимаемых местообитаний) приводит к формированию подобных демонстрационных признаков, выглядящих своего рода излишествами (достаточно вспомнить хотя бы хвост павлина или птицы-лиры, ослепительные окраски фазанов и райских птиц и т.п.).
Из плейстоценовых отложений известны многочисленные находки крупных нелетающих страусоподобных птиц, большинство видов которых вымерло к концу плейстоцена или были истреблены человеком в совсем недавнее время. Эта судьба постигла в первую очередь крупных птиц, обитавших на островах: мадага-скарских эпиорнисов (Aepiornithes), новозеландских моа (Dinor-nithes) и др.
Как уже упоминалось, прогрессировавшее похолодание завершилось в плейстоцене развитием великого оледенения. Наступление ледников сопровождалось смещением климатических зон к экватору, вместе с которыми отступали теплолюбивые фауны и флоры, отчасти подвергавшиеся при этом вымиранию. В межледниковые эпохи, продолжавшиеся по нескольку десятков тысяч лет, происходили обратные смещения климатических зон и соответствующих флор и фаун.
В то же время в относительной близости к границам оледенения в Евразии и Северной Америке сформировался специфический приледниковый пояс с особыми физико-географическими условиями: резко континентальный климат с низким уровнем средних температур при сухом воздухе и значительной обводненности территории летом за счет талых ледниковых вод, с возникновением в низинах озер и болот. В этой обширной приледниковой зоне возник особый биоценоз - тундростепь, существовавшая все время оледенения и перемещавшаяся в соответствии с изменениями границ ледника к северу или к югу. Флора тундростепи включала различные травянистые растения (особенно злаки и осоки), мхи, а также мелкие деревца и кустарники, произраставшие главным образом в долинах рек и по берегам озер: ивы, березы, ольхи, сосны и лиственницы. При этом общая биомасса растительности в тундростепи была, по-видимому, очень велика, в основном за счет трав, что позволило расселиться на огромных пространствах приледникового пояса обильной и своеобразной фауне, которую называют мамонтовой.
Эта удивительная приледниковая фауна включала мамонтов, шерстистых носорогов, овцебыков, короткорогих бизонов, яков, северных оленей, антилоп-сайгаков и дзеренов, лошадей, куланов, грызунов - сусликов, сурков, леммингов, зайцеобразных, а также различных хищников: пещерных львов, пещерных медведи, волков, гиен, песцов, росомах. Состав мамонтовой фауны свидетельствует о том, что она произошла от гиппарионовой фауны, будучи ее северным приледниковым вариантом, тогда как современная африканская фауна является южным, тропическим производным гиппарионовой.
Для всех животных мамонтовой фауны характерны приспособления к жизни в условиях низких температур, в частности длинная и густая шерсть. Густой и очень длинной рыжей шерстью с длиной волос до 70-80 см был покрыт и мамонт (Mammonteus, рис. 93) - северный слон, обитавший 50-10 тыс. лет назад на огромных территориях Европы, Азии и Северной Америки.

Изучение представителей мамонтовой фауны значительно облегчается сохранением целых трупов или их частей в условиях вечной мерзлоты. На территории нашей страны был сделан целый ряд замечательных находок такого рода. Наиболее известен из них так называемый "березовский" мамонт, найденный в 1901г. на берегу реки Березовки в Северо-Восточной Сибири, а последняя находка - почти целый труп мамонтенка 5-7 месяцев от роду, обнаруженный в 1977г. на берегу ручья, впадающего в реку Берелех (приток Колымы).
По пропорциям тела мамонт заметно отличался от современных слонов, индийского и африканского. Теменная часть головы сильно выступала вверх, а затылок был скошен вниз, к глубокой шейной выемке, позади которой на спине возвышался большой горб, состоявший из жира. Вероятно, это был запас питательных веществ, использовавшийся во время голодного зимнего сезона. Позади горба спина была круто скошена вниз. Огромные бивни, до 2,5 м длины, закручивались вверх и внутрь. В содержимом желудков мамонтов были обнаружены остатки листьев и стеблей злаков и осок, а также побегов ив, берез и ольх, иногда даже лиственниц и сосен. Основу питания мамонта, вероятно, составляли травянистые растения.
Во многих местах, где прежде обитали мамонты: в Сибири, на Новосибирских островах, на Аляске, на Украине и др. - были обнаружены огромные скопления скелетов этих животных, так называемые "Мамонтовы кладбища". О причинах возникновения мамонтовых кладбищ было высказано немало предположений. Наиболее вероятно, что они сформировались, как и большинство массовых скоплений ископаемых остатков наземных животных, в результате сноса течением рек, особенно во время весенних половодий или летних разливов, в разного рода естественные отстойники (заводи, омуты, старицы, устья оврагов и т.п.), где целые скелеты и их фрагменты накапливались в течение многих лет.
Вместе с мамонтами обитали шерстистые носороги (Coelodonta), покрытые густой коричневой шерстью. Облик этих двурогих носорогов, так же как мамонтов и других животных этой фауны, был запечатлен людьми каменного века - кроманьонцами в их рисунках на стенах пещер. На основании археологических данных можно с уверенностью утверждать, что древние люди охотились на самых различных животных мамонтовой фауны, в том числе и на шерстистых носорогов и самих мамонтов (а в Америке на еще сохранявшихся там мастодонтов и мегатериев). В связи с этим высказывались предположения, что человек мог сыграть определенную роль (по мнению некоторых авторов - даже решающую) в вымирании многих плейстоценовых животных.
Вымирание мамонтовой фауны отчетливо коррелирует с концом последнего оледенения 10-12 тыс. лет назад. Потепление климата и таяние ледников резко изменили природную обстановку в прежнем поясе приледниковой тундростепи: значительно увеличились влажность воздуха и выпадение осадков, как следствие, на больших территориях развилась заболоченность, зимой возросла высота снегового покрова. Животные мамонтовой фауны, хорошо защищенные от сухого холода и способные добывать себе пищу на просторах тундростепи в малоснежные зимы ледниковой эпохи, оказались в крайне неблагоприятной для них экологической обстановке. Обилие снега зимой сделало невозможным добывание пищи в достаточном количестве. Летом же высокая влажность и заболачивание почвы, крайне неблагоприятные и сами по себе, сопровождались колоссальным возрастанием численности кровососущих насекомых (гнуса, столь обильного и в современной тундре), укусы которых изнуряли животных, не давая им спокойно Кормиться, как это происходит и ныне с северными оленями. Таким образом, мамонтовая фауна оказалась за очень короткий срок (таяние ледников происходило очень быстро) перед лицом резких изменений среды обитания, к которым большинство составлявших ее видов не смогло столь быстро приспособиться, и мамонтовая фауна как целое прекратила свое существование. Из числа крупных млекопитающих этой фауны сохранились до наших дней северные олени (Rangifer), обладающие большой подвижностью и способные совершать дальние миграции: летом в тундру к морю, где меньше гнуса, а на зиму на ягельные пастбища в лесотундру и тайгу. В относительно малоснежных местообитаниях на севере Гренландии и на некоторых островах Североамериканского архипелага сохранились овцебыки (Ovibos). Приспособились к новым условиям некоторые мелкие животные из состава мамонтовой фауны (лемминги, песцы). Но большинство видов млекопитающих этой замечательной фауны вымерли к началу голоценовой эпохи.
(По некоторым данным, в голоцене 4-7 тыс. лет назад на острове Врангеля еще сохранялась популяция измельчавших мамонтов) (См. кн.: Верещагин Н.К. Почему вымерли мамонты. - М.. 1979).
В конце плейстоцена произошло еще одно значительное изменение фауны, правда ограниченное территорией Америки, но остающееся до сих пор загадочным. В обеих Америках вымерло подавляющее большинство крупных животных, столь обильных там прежде: и представители мамонтовой фауны, и обитавшие в более южных районах, где не было никакого оледенения, мастодонты и слоны, все лошади и большинство верблюдов, мегатерии и глиптодонты. Видимо, еще в плиоцене исчезли носороги. Из крупных млекопитающих сохранились лишь олени и бизоны в Северной Америке и ламы и тапиры в Южной. Это тем более удивительно, что Северная Америка была родиной и центром эволюции лошадей и верблюдов, сохранившихся до нашего времени в Старом Свете.
Нет никаких признаков значительных изменений условий существования в конце плейстоцена на большей части территории Америки, не подвергавшейся оледенению. Более того, после появления в Америке европейцев некоторые из завезенных ими лошадей одичали и дали начало мустангам, быстро размножившимся в северо-американских прериях, условия которых оказались благоприятными для лошадей. Индейские племена, жившие охотой, не оказывали значительного влияния на численность огромных стад бизонов (и мустангов после их появления в Америке). Человек на уровне культуры каменного века едва ли мог сыграть решающую роль в вымирании многочисленных видов крупных плейстоценовых животных (за исключением, может быть, медлительных и малосообразительных мегатериев) на обширных территориях обеих Америк.
После завершения последнего оледенения 10-12 тыс. лет назад Земля вступила в голоценовую эпоху четвертичного периода, в течение которой установился современный облик фауны и флоры. Условия жизни на Земле ныне значительно более суровы, чем на протяжении мезозоя, палеогена и большей части неогена. И богатство и разнообразие мира организмов в наше время, судя по всему, существенно ниже, чем за многие прошлые геологические эпохи.
В голоцене все сильнее проявляется воздействие человека на среду обитания. В наше время с развитием технической цивилизации деятельность людей стала поистине важнейшим глобальным фактором, активно, хотя в большинстве случаев непродуманно и губительно, изменяющим биосферу.
В связи с формированием человека современного вида (Homo sapiens) и развитием человеческого общества в течение четвертичного периода А.П.Павлов предложил назвать этот период кайнозойской эры "антропогеном". Обратимся же теперь к эволюции самого человека.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 1261; Нарушение авторских прав?; Мы поможем в написании вашей работы!