КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
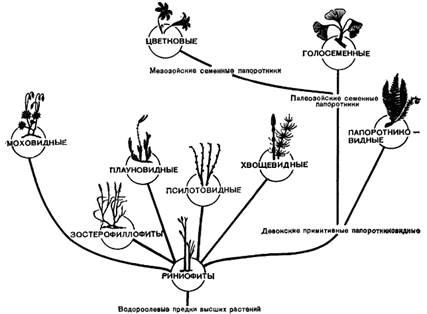
Систематика высших растений
|
|
|
|
Проблема происхождения высших растений
Предпосылки выхода растений на сушу. Появление наземных, или высших, растений ознаменовало начало новой эры в жизни нашей планеты. Освоение растениями суши сопровождалось появлением новых, наземных, форм животных; сопряженная эволюция растений и животных привела к колоссальному разнообразию жизни на земле, изменила ее облик.
Предпосылки появления наземных растений:
1. независимый ход эволюции растительного мира подготовил появление новых, более совершенных форм.
2. за счет фотосинтеза морских водорослей в атмосфере земли произошло увеличение количества кислорода; к началу силурийского периода оно достигло такой концентрации, при которой оказалась возможной жизнь на суше.
3. в начале палеозойской эры на обширных территориях земли происходили крупнейшие горообразовательные процессы, в результате которых возникли Скандинавские горы, горы Тянь-Шань, Саяны. Это вызвало обмеление многих морей и постепенное появление суши на месте бывших мелких водоемов. Если раньше водоросли, населявшие литоральную зону, только в отдельные кратковременные периоды жизни оказывались вне воды, то по мере обмеления морей они переходили к более длительному пребыванию на суше. Это, очевидно, сопровождалось массовой гибелью водорослей; выживали лишь те немногие растения, которые смогли противостоять новым условиям жизни.
Выход растений на сушу начался, очевидно, в конце силура (400 млн. лет назад).
В 1859 году канадский геолог Джеймс Досон в девонских отложениях на полуострове Гаспе в Канаде обнаружил остатки удивительно примитивного высшего растения не похожего ни на одно из известных науке в то время. Его вильчато разветвленные стебли были лишены листьев, а проводящая система имела примитивный тип организации. Досон дал своему растению название Psilophyton princeps, что можно перевести на русский язык как "голорос первичный".
|
|
|
Позднее в Шотландии были найдены и другие, еще более примитивные растения: риния, хорнеофит и куксония. Эти древние растения были выделены в особый отдел, для обозначения которого прежде употреблялось название "псилофиты" (Psilophyta), но в настоящее время многие ученые предпочитают название "риниофиты" (Rhiniophyta).
Риниофиты – это силуро-девонские растения, которые дали начало всему разнообразию наземной флоры, относятся к пойкилогидричесим растениям. Они жили наполовину в воде, водный обмен у них не стабилизирован и жизненные процессы зависели от наличия влаги в окружающей среде. Предполагается, что предками всех существующих растений были водоросли, в частности, зеленые.

Первые наземные растения стелились по земле, тело было дорзовентральной («дорзум» - спина, «вентер» - живот) структуры пластинчатой формы, с воздухом соприкасались одной стороной.
С момента выхода на сушу растения развиваются в двух основных направлениях: гаметофитном и спорофитном. Высшим растениям свойственна правильная смена поколений в цикле их развития. Растение имеет две фазы развития, которые сменяют одна другую: гаметофит и спорофит. Гаметофит — это половое поколение, на котором образуются половые органы — антеридии и архегонии. Спорофит — неполовое поколение, на котором формируются органы неполового размножения. Спорофит — это нормально развитое растение, которое имеет корень, стебель и листья. На спорофите образуются споры, которые прорастают и дают начало гаметофиту. Подобная смена поколений в цикле развития растений сложилась эволюционно, в ходе естественного отбора. Гаметофитное направление было представлено мохообразными, а спорофитное — остальными высшими растениями, включая цветковые. Спорофитная ветвь оказалась более приспособленной к наземным условиям.
|
|
|
Таким образом, как приспособление к новым условиям жизни на суше появляется спорофит, тело которого приобрело стеблевидную форму. Это был небольшой стебель с верхушечной точкой роста и вертикальным положением, зеленый. Впоследствии как приспособление к увеличению ассимилирующей поверхности появляются листья – вторичные органы и последним возникает корень.
Предки современных наземных растений имели форму дихотомически ветвящихся теломов. В результате процесса «перевершинивания» происходит смена типа ветвления и морфологическая и физиологическая дифференциация ветвей. Более сильные ветви, принявшие направление материнских, становятся осевым органом (стеблем) с осевой симметрией. Более слабые ветви прекращают верхушечный рост, прекращается ветвление и они уплощаются, становятся листьями.
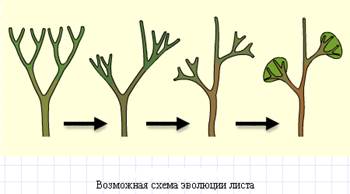
Происхождение стебля
Стебель — существенная часть растений, он легко воспроизводит и корни, и листья. Главные функции стебля — механическая поддержка кроны и проводящая. По коре его идет ток питательных веществ из листьев в корни, чем обусловливается рост и корневой системы. По древесине, идет водный ток из корней в листья. Основная форма стебля — форма колонны — так проста, что в этом отношении никакой особой эволюции от крупной морской водоросли лессонии через стволы плауновых деревьев каменноугольной эпохи к стволам крупных хвойных и лиственных деревьев не замечается. Наоборот, анатомия стебля дает очень сложную картину постепенного усложнения и усовершенствования проводящего аппарата.
У бурых водорослей в центральной части их стебля находятся длинные трубчатые клетки, имеющие анастомозы и сообщающиеся одна с другой с помощью ситовидных пластинок. Последнее — не что иное, как поперечные перегородки, отделяющие одну клетку от другой. Протоплазмы же их проходят через поры сит, неся с собой питательные вещества. Ствол массивен, ассимиляция происходит в поверхностных слоях ткани, внутренние части, затененные наружными, будут голодать, если их не будет пронизывать система трубочек с движущимися растворами внутри.
|
|
|
У мхов, в их тонких стеблях, в центре находится проводящий цилиндр из узких тонких трубчатых клеток, образующих точно пригнанные вертикальные ряды.
Поперечные разрезы шлифов стеблей псилофитов показывают сразу, что эти первые растения суши были построены сложнее мхов, хотя и сходны с ними. У ринии в центре стебля находился участок толстостенных клеток, проводивших воду от корневища к верхушкам побегов. Участок этот был окружен кольцом многочисленных тонкостенных трубчатых клеток, проводивших питательные вещества от зеленых верхних частей растения в корневище.
|

У астероксилона на продольных шлифах видны водоносные клетки — трахеиды с характерными кольчатыми или сетчатыми утолщениями на стенках.
|
|


Совокупность всех таких проводящих клеток, имеющая вид внутреннего шнура, пролегающего между более рыхлой мякотью коры стебля, получила наименование стеле, или столба.
Эволюция стеле папоротников, приведшая к большому разнообразию и строения, и расположения, в тех случаях, когда их много, привела в конце концов к выработке наиболее рационального построения древесины и коры у цветковых растений.
Если у папоротников древесина всегда состоит из водоносных клеток или трахеид, то у хвойных в их молодых тканях уже есть небольшие спиральные сосуды (в протоксилеме), у гнетовых их значительно больше и они принимают некоторое участие также и в строении вторичной древесины. У однодольных не развит еще камбий, т. е. специальная зародышевая или образовательная ткань проводящих пучков, что мешает им срастаться вместе и расти в толщину. Поэтому у этих строение стволов менее плановое, менее совершенное, чем у двудольных растений, дающих благодаря камбию полное разделение ствола на кору и древесину.
|
|
|
Таким образом, процесс эволюции охватывает все детали строения стебля, делая его достаточно совершенным орудием как механической крепости растения, так и физиологической связи между двумя активными системами органов растительной жизни, именно между листовой и корневой системами.
Происхождение листа
Первые листья, которые вполне оправдывают такое их обозначение, — это листья некоторых псилофитов (род Asteroxylon), обладавшие, по-видимому, уже вполне развитым устьичным аппаратом. Листья ископаемых и современных плаунов следуют за ними. Эти листья все еще примитивные: у них нет завершенного разделения на черешок и пластинку, нет развитой сети жилок; анатомически оба основных аппарата листа, фотосинтетический — палисадная паренхима — и транспирационный — паренхима губчатая, выражены несовершенно, как несовершенно и различие между верхней и нижней сторонами листа. Правда, подобные листья встречаются и у растений, принадлежащих к высоко организованным семействам, но там это результат упрощения, вызываемого внешними условиями, здесь же — первичное строение.
У папоротникообразных, названных Джеффреем Pteropsida, листья крупные, с хорошей дифференцировкой на черешок и пластинку. Их примитивность сказывается главным образом в там, что они растут верхушкою, а не основанием, как листья цветковых. Такой способ роста позволяет им развивать нередко добавочный рост отдельных частей листа, курчавость краев и другие уродливости.
У цветковых имеется колоссальное разнообразие листовых форм и строений. Сеть жилок в своей более простой форме дает очень мало анастомозов. Главные жилки располагаются параллельно одна другой, и при перерезке любой из них примыкающий к верхней части перерезанной жилки участок ткани засыхает. Более прогрессивный тип жилкования — сетчатый. Если перерезать одну из главных жилок у листа этого типа, то ткань кругом остается жива, так как получит воду обходным путем, через боковую сеть.
Такие сложные листья, как листья гороха, акации и массы других бобовых, со свободным движением отдельных частей, с организованным отводом продуктов ассимиляции из тканей (мякоти) в ситовидные трубки и пр., являются наиболее совершенным выражением эволюции листа. От листа плауновых до листа гороха пройден сложный и долгий путь, приведший к выработке прекрасного пластичного, сообразно условиям среды, аппарата фотосинтеза; фотосинтез же, как известно, — это главный физиологический процесс зеленых растений.
Происхождение корня
Полярность, т. е. противоположение испаряющей части растения и части, всасывающей воду, — вот первый импульс к образованию первых корнеподобных органов.
Гаметофиты мхов, даже наиболее крупных (исключение - сфагновые мхи и мхи подводные), поглощают воду ризоидами, т. е. отдельными клетками, покрывающими нижнюю часть стебля. Никакого подобия настоящего корня у мхов нет, как вообще его нет ни у одного гаметофита гаплоида.
У древнейшей группы наземных растений, у псилофитов, корней не было вовсе, воду они вбирали, так же как и мхи, с помощью ризоидов. Только у рода астероксилон, побеги которого слабо напоминают плауновые растения, есть вильчато разветвленные, отходящие от корневища отростки, напоминающие корни.
У плаунов, хвощей и у всех папоротников, как у растений диплоидных, корни уже вполне развиты. Они всегда невелики, отходят от корневища или от коры стебля и неглубоко погружаются в почву. Ввиду того, что все эти растения жили и живут во влажных местностях, они могут довольствоваться поверхностными корнями.
Чем крупнее растение, чем более развита его испаряющая крона, тем более повышается потребность в глубоко зарывающейся в почву мощной корневой системе.
У голосеменных растений впервые появляется стержневой корень, закладывающийся уже в период развития зародыша в семени. Такой корень дает возможность молодому растению быстро углубиться в почву и распределить боковые корни в таком слое почвы, который наилучше обеспечивает водоснабжение растения.
Однако у корня есть и еще функция — это функция прикрепления к почве, функция механическая. Уже у крупных морских водорослей типа ламинарии от нижней части стебля отходят мощные ризоиды, впивающиеся в камни и удерживающие водоросль на определенной глубин. Здесь функция всасывания не имеет значения, все дело в механическом сопротивлении среде.
Корни наших деревьев должны отвечать огромному сопротивлению, так как ветер очень сильно давит на обширную площадь кроны; дерево гнется, иногда ломается, а корни не поддаются, сохраняя связь между деревом и почвой. Зато при условиях, неблагоприятных развитию корневой системы, ветровал неизбежен.
У растений, живущих на очень сухих почвах, поражает длина корней, часто во много раз превышающая в вышину наземные части растения.
Анатомически корень состоит из основного цилиндра, коры, конуса нарастания и защищающего этот конус чехлика. Кора на некотором расстоянии от конца корня несет зону корневых волосков, усиливающих всасывание. Осевой цилиндр содержит в себе проводящие воду сосуды, через которые она поступает в стебель. Клетки коры развивают колоссальное осмотическое давление, обеспечивающее движение воды и сосудах. Если корень находится в симбиозе с грибками, образуя микоризу, то корневые волоски, как правило, отсутствуют.
Наконец, корень постоянно растет и своими движениями при этом роет землю. Подвергаясь действию внешних факторов: силе тяжести, свету, теплу, влажности, присутствию кислорода воздуха и химизму почвенного раствора, — корень растет неравномерно, искривляется и проникает в наиболее благоприятные для него слои почвы. Таков результат длительной, постоянно поддерживающейся борьбы за существование. Разумеется, и эта способность корня ориентироваться в почве далась не сразу, а выработалась постепенно.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 2242; Нарушение авторских прав?; Мы поможем в написании вашей работы!