КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Резерв наследственной изменчивости вида и его мобилизация
|
|
|
|
Взаимодействие всех разобранных процессов — постоянного мутирования, элиминации вредных и распространения безвредных или условно и частично вредных мутаций через процесс размножения, их разнообразного комбинирования в результате свободного скрещивания (панмиксии), под непрерывным контролем естественного отбора, поощряющего распространение наиболее благоприятных комбинаций, и закрепления некоторых комбинаций в условиях локального или временного ограничения панмиксии — ведет к накоплению внутри любой популяции известного запаса индивидуальной изменчивости. Такой внутривидовой запас изменчивости определяет эволюционную пластичность данной популяции при изменении соотношений между организмом и факторами внешней среды. Как видно, это накопление резерва изменчивости зависит от многих факторов: скорости мутирования, остроты элиминации и интенсивности естественного отбора, а также от свободы комбинирования, зависящей, в свою очередь, от величины популяции, степени ее изолированности и т. д. Поэтому в различных условиях борьбы за существование и эволюционная пластичность разных популяций и разных видов организмов бывает весьма различной.
А. УСЛОВИЯ НАКОПЛЕНИЯ РЕЗЕРВА ИЗМЕНЧИВОСТИ (В ПОПУЛЯЦИЯХ)
Значение мутабилъности. Лишь при непрерывном появлении все новых мутаций они могут накапливаться в популяции. Единичные мутации обычно теряются, даже и в том случае, если они благоприятны для данного вида организмов в данных условиях. С другой стороны, повторно возникающие мутации могут при достаточно высоком мутационном «давлении» накапливаться в популяции до известной степени и в том случае, если они неблагоприятны. Они могут преодолеть «давление» отбора вплоть до достижения уровня равновесия, определяемого интенсивнос-тями обоих противоположных процессов.
|
|
|
Существуют как высоко мутабильные популяции и линии, так и мало мутабильные. По Р. Л. Берг, мутабильность обыкновенной дрозофилы (Drosophila melanogaster) из больших популяций Умани и Никитского сада в Крыму очень высока; мутабильность той же дрозофилы из малых популяций Дилижана (Армения) и Каширы (Московская область) очень низка. Низка также мутабильность американской линии «Флорида». Однако и в популяциях с низкой мутабильностью встречаются отдельные очень высоко мутабильные линии (Demerec, Шапиро, Берг).
Встречаются и мутации, увеличивающие темп мутирования. Мутабильность оказывается таким же наследственным признаком, меняющимся в процессе мутирования, как и любые другие признаки организма. Генетики говорят и о «генах» мутабилъности, так же как о «летальных генах» или о «генах» стерильности. Само собой разумеется, что это лишь упрощенный способ выражения той мысли, что некоторые наследственные изменения одного «гена» вносят столь значительное расстройство в координи-рованность морфофизиологических (включая биохимические) процессов, что делают организм нежизнеспособным, делают невозможным его развитие (летали), прекращают развитие половых желез или половых клеток (гены «стерильности»). Иногда же эти изменения лишь немного нарушают «баланс» наследственного аппарата так, что он оказывается менее устойчивым и в нем легче возникают новые мутации (гены «мутабилъности»).
Значение темпа мутирования для скорости распространения мутаций в популяции ясно само собой. Для нас важно, что при прочих равных условиях, т. е. прежде всего при равной интенсивности элиминации, повышенная частота возникновения мутаций («мутационное давление») ведет к увеличению концентрации мутаций и к установлению равновесия на более высоком ее уровне. Таким образом, в высоко мутабильных популяциях и насыщенность их мутациями должна достигать более высокого уровня. Мы уже видели, что она для неблагоприятных мутаций достигает равновесия на уровне, пропорциональном частоте мутирования (и) и обратно пропорциональном коэффициенту селекции (s) в пользу нормы, согласно формуле для нормы q — 1 — u/s, а следовательно — для мутации: (1 — q — u/s).
|
|
|
Изменение мутабильности является, очевидно, лишь косвенным результатом эволюции. При обострении борьбы за существование и увеличении интенсивности избирательной элиминации, — особенно в относительно постоянных и однородных условиях внешней среды, —- всех более значительных уклонений от приспособленной нормы (стабилизирующая форма отбора) происходит сокращение изменчивости, т. е. «иммобилизация» вида. Косвенным результатом элиминации большого числа мутаций является преобладающая элиминация мутабильных линий, как дающих максимум мутаций, и, следовательно, преобладающее переживание более однородного («нормального») потомства менее мутабильных линий.
Обратный процесс повышения мутабильности может быть результатом сохранения вновь возникающих линий с более высокой мутабильностью (мутации «мутабильности») в силу ослабления интенсивности элиминации, при уменьшении остроты борьбы за существование, особенно в условиях разнообразной и меняющейся внешней среды. Возникновение таких линий в результате отдельных мутаций наблюдалось неоднократно (Н. Шапиро, 1938). Простое их сохранение при благоприятных условиях ведет к увеличению мутабильности популяций через распространение этих мутаций до нового уровня равновесия (определяемого хотя и косвенно, но в конце концов также через соотношение u/s).
В некоторых случаях повышение мутабильности может быть результатом и другого процесса. В межгрупповой конкуренции при меняющихся условиях среды побеждают те популяции, которые быстрее приспосабливаются к новым условиям существования. При прочих равных условиях побеждать будут более мута-бильные популяции и более мутабильные линии. Таким образом, мутабильность может меняться в результате межгруппового соревнования в темпах эволюции (Р. Берг). Не отрицая значения межгруппового соревнования в определении мутабильности, следует отметить, что указанный ранее фактор — интенсивность элиминации мутаций, а следовательно и мутабильных линий — должен быть значительно более эффективным. К тому же он независим от темпов эволюции и скорости изменения внешней среды. Интенсивность элиминации определяется лишь условиями борьбы за существование, и это вполне объясняет наличие существующих в настоящее время различий. В однородных условиях существования малых популяций преобладает иммобилизирующая роль отбора (острая элиминация уклонений) и, следовательно, преимущественное распространение менее мутабильных линий. В разнородных и благоприятных условиях существования больших популяций, с возможностью широких миграций, элиминация уклонений не столь остра и вполне возможно общее повышение уровня изменчивости, а вместе с тем и сохранение более мутабильных линий.
|
|
|
Б. СОСТАВ МОБИЛИЗАЦИОННОГО РЕЗЕРВА
Вредные мутации и комбинации постоянно элиминируются, а полезные быстро завоевывают всю популяцию. Поэтому мобилизационный резерв видов и популяций составляется в основном из «нейтральных» мутаций и «сбалансированных», т. е. из безразличных и, быть может, частично благоприятных их комбинаций. Вопрос о самом существовании вполне нейтральных мутаций подвергается справедливому сомнению. Что же касается неблагоприятных мутаций, то они, казалось бы, и не могут накапливаться. Не нужно, однако, подходить слишком формально к пониманию «нейтральности» мутаций или их комбинаций. К нейтральным мутациям следует причислить все мутации, которые не получают заметного выражения вследствие наличия совершенного регуля-торного аппарата, сглаживающего нарушенные соотношения.
К категории нейтральных мутаций можно отнести прежде всего вообще все малые мутации, обладающие столь незначительным выражением, что они не могут обусловить заметного снижения жизнеспособности их обладателей. Предполагается, что малые мутации возникают чаще. Однако, если это даже и неверно, то насыщенность природных популяций малыми мутациями, во всяком случае, очень высока. Многообразие выражения многих мутаций, которое не только в разных расах, популяциях и линиях (Тимофеев-Ресовский, Добжанский, Дубинин), но даже у отдельных особей оказывается различным; неполнота проявления многих мутаций даже в строго контролируемых идентичных условиях культуры,— все это указывает на существование многих небольших, трудно анализируемых, но именно генотипических различий между популяциями, между разными линиями и даже отдельными особями наших обычных культур. Экспериментально это может быть продемонстрировано внесением определенной мутации (например, beaded) в различные линии или успег.шыми результатами искусственного отбора на максимальное или минимальное выражение известного признака (Камшилов, 1939; Шифрин, 1941). В особенности изменчиво выражение многих мутаций в гетерозиготном состоянии. Быстрота, с какой меняется это выражение в лабораторных линиях, показывает высокий уровень насыщения малыми мутациями даже чистых культур, во всяком случае — численно очень ограниченных, экспериментальных популяций. Значение малых мутаций в эволюции также бесспорно.
|
|
|
Во-вторых, к категории нейтральных мутаций можно отнести и более значительные мутации, если они в гетерозиготном состоянии заметно не нарушают общей жизнеспособности организма. Это касается главным образом рецессивных и полудоминантных мутаций. Так как первоначальное накопление мутаций касается всегда гетерозиготы, то все рецессивные (даже вредные) мутации распространяются совершенно свободно. Однако новые мутации, как правило, полудоминантны (Фишер, 1931) и, следовательно, имеют некоторое выражение и в гетерозиготе. Если это выражение не слишком неблагоприятно, по крайней мере в некоторых генотипах, то и такие мутации могут накапливаться. В процессе их накопления и комбинирования они могут потерять свои вредные выражения (через отбор малых мутаций — модификаторов). Природные популяции дрозофилы действительно насыщены и вредными мутациями не только рецессивными, но и полудоминантными, выражения которых могут быть подавлены модификаторами, имеющими частично эпистатическое влияние (Берг, Гершензон).
Можно считать доказанным, что в состав резерва наследственной изменчивости входят и более крупные мутации, вредные выражения которых частью погашены в процессе подбора модификаторов. Это — большая категория нейтрализованных или обезвреженных мутаций, которые по мере их нейтрализации могут достигнуть все более высокого уровня насыщения популяции. В этом случае, как и в предыдущем, популяция может быть по внешности весьма однородной. Индивидуальные различия очень невелики, так как выражения отдельных мутаций в значительной мере погашены.
В-третьих, мы можем условно причислить к той же категории нейтральных мутаций, входящих в состав мобилизационного запаса, и частично вредные (частично полезные) мутации, которые, будучи в некоторых отношениях неблагоприятными, оказываются в других отношениях (на других стадиях или в других своих выражениях) благоприятными. Такие мутации для организма в целом оказываются действительно нейтральными. В процессе эволюции вредные выражения таких мутаций будут погашаться (подбором модификаторов) и накопление их может идти возрастающими темпами по мере того, как они становятся все более благоприятными.
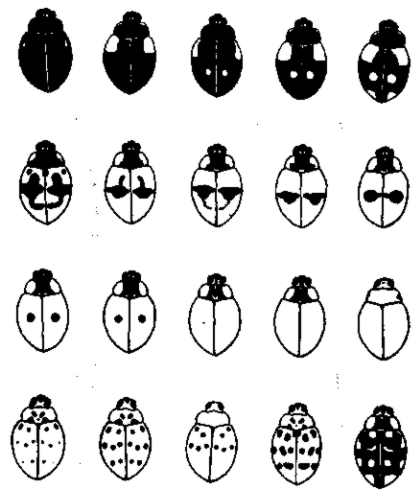
Рис. 18. Гетероморфизм окраски у божьей коровки Adalia bipunctata. Изображены только гомозиготные формы. По Я. Я. Лусу, 1932
В-четвертых, к тем же нейтральным мутациям мы отнесем и мутации условно вредные (условно полезные). Если популяция живет в разнородной среде, то локальные условия существования предъявляют различные требования к организму. Организм, вполне приспособленный к одним локальным условиям, оказывается менее приспособленным к другим локальным условиям. Мутация, неблагоприятная в одних условиях (например, мелани-стическая форма на светлом фоне), может оказаться благоприятной в других (та же меланистическая форма на темном фоне).
Гетероморфизм. При наличии известного, более или менее постоянного, разнообразия условий, характерного для данного ареала распространения, возможно установление локального, более или менее постоянного соотношения разных форм. Одни и те же наследственные формы входят в состав различных популяций данного вида в различном процентном соотношении. Таково, например, географическое распределение разных типов окраски у божьих коровок (Harmonia oxyridis, по Добжанскому, 1937, и другие виды — рис. 10, 18). Подобным же образом весьма постоянное соотношение имеется между разными типами полосатых и гладких (т. е. однотонных) раковин садовых улиток (Cepea hortensis и С. nemoralis). Это соотношение мало меняется даже в течение геологических периодов (Diver, 1929, по Холдену, 1935). Такую форму генетического полиморфизма можно обозначить термином равновесный гетероморфизм. Еще ярче выражен такой гетероморфизм у обыкновенного богомола (Mantis religiosa). Он характеризуется одновременным существованием различных ясно выраженных защитных окрасок (рецессивной — зеленой, и доминантной — буро-желтой) у отдельных особей в известном числовом соотношении (для Карадага в Крыму 1:2,5), что связано с их распределением на соответствующей растительности (Беляев, 1946). Зеленые держатся на зеленых растениях (в Карадаге на каперцах), а буро-желтые — на высохших. Подобный же равновесный гетероморфизм ограничивается иногда одним полом, как в случае мимикрии у самок Papilio dardanus и P. polytes, генетический механизм которого разобран Фишером (1930).
Внутривидовой гетероморфизм имеет приспособительное значение. Каждая форма в целом все полнее приспособляется к известным частным условиям существования. Таким образом, возможна дальнейшая дифференциация вида на локальные формы (экотипы). Для разбираемых здесь вопросов важно, что в случае изменения внешней среды (ее климатических или биотических факторов) условия, бывшие прежде случайными или локальными (засуха, холод, недостаток света, влажность, размножение хищников или организмов, служащих пищевым материалом), могут стать более частыми, постоянными или всеобщими. В ответ на такое изменение в данной популяции произойдет в основном лишь сдвиг в сторону преобладания одной из ранее существовавших форм, которая теперь получила постоянное преимущество. Малочисленная второстепенная форма может быстро стать преобладающей. В таких случаях можно говорить об известной степени «пре-адаптации» данного вида к новым условиям в некотором числе особей. Такая преадаптация существует, однако, только благодаря тому, что эти новые условия встречались уже и раньше в жизни вида, но не как постоянные, а как случайные, периодические или локальные особенности существования.
Несомненно, что известный гетероморфизм, постоянно наблюдаемый в популяциях вида, живущего в разнообразных условиях внешней среды, значительно повышает его эволюционную пластичность. Возникая на базе индифферентного мобилизационного резерва мутаций, накапливающихся в популяции и комбинируемых с тем большей скоростью, чем многочисленнее популяция и чем более разнообразны условия внешней среды в пределах ареала ее распространения, этот гетероморфизм еще до распада вида на экотипы и географические расы начинает приобретать приспособительное значение в известных условиях существования. В этом случае довольно быстрое изменение в факторах внешней среды может привести к очень быстрой мобилизации внутренних ресурсов и быстрому преобразованию вида в процессе эволюции. Чем больше запас индивидуальной изменчивости и генетический полиморфизм популяций, тем выше эволюционная пластичность вида.
Обычный резерв индивидуальной изменчивости имеет в моно-морфном виде в значительной мере скрытый характер — по внешности особи могут быть весьма однородными («дикий» тип дрозофилы) даже и в больших популяциях.
В. УСЛОВИЯ ВСКРЫТИЯ И МОБИЛИЗАЦИИ РЕЗЕРВОВ
Мобильность видов означает их потенциальную способность к эволюции. Эта потенциальная способность может быть реально использована лишь в известных условиях. Естественный отбор имеет полем своего приложения всегда лишь реализованные фенотипы, а не потенциально возможные. Поэтому непосредственным материалом для движущей формы отбора и для прогрессивной эволюции могут быть лишь мутации и их комбинации, имеющие какое-либо физиологическое или морфологическое выражение, а не скрытые формы мутаций. Лишь явная наследственная гетерогенность популяций, т. е. наличие индивидуальных уклонений или зарождающихся форм групповой изменяемости (гетероморфизм), может быть непосредственно использована в эволюции организмов. Все отрегулированные или нейтрализованные формы изменчивости, входящие в состав «мобилизационного резерва», должны быть сначала выявлены, мобилизованы и лишь после этого могут стать предметом прямого отбора (в его ведущей форме). Таким образом, первым ответом вида на изменение внешней среды, или точнее — на изменение соотношений между организмом и внешней средой, является более или менее быстрое вскрытие его резервов изменчивости.
а. Вскрытие резервов изменчивости. Вскрытие резервов осуществляется прежде всего в процессе гомозиготизации мутаций и их комбинаций, которые в новых условиях существования оказываются благоприятными. В более широком масштабе это возможно при разбивке вида на малые изолированные популяции, разбивке, ведущей к родственным скрещиваниям. Гомозиготиза-ция может принять всеобщий характер при переходе к апогамии, что наблюдается нередко у растений. В этом случае очень ясно проявляется гетерогенность популяций (одуванчик, ястребинка, крупка и др.).
Второй путь лежит через индивидуальное приспособление и снятие в связи с этим некоторых регуляций. Изменение внешних факторов может и прямо коснуться регуляторного аппарата, всегда отличающегося известной лабильностью, и несколько нарушить полноту регуляции. С другой стороны, еще важнее, что в процессе индивидуального приспособления меняются функциональные соотношения частей, т. е. теряют свое значение некоторые прежние соотношения и устанавливаются новые. Прежняя «регуляция» теряет свою координированность и снимается в том же порядке индивидуального взаимоприспособления частей, в каком она в свое время возникла. Это будет способствовать выявлению таких наследственных изменений, которые ранее сглаживались путем различных функциональных компенсаций. В новых условиях эти компенсации просто отпадают.
Изменение внешних факторов может и непосредственно повлиять на выражение существующих мутаций. Это выражение было погашено лишь в прежних условиях развития организма и может так или иначе проявиться в новых условиях (мы знаем, что выражение мутации зависит как от генотипа организма, так и от внутренних факторов его развития и, наконец, и от факторов внешней среды, см. 1–4). При воздействии внешних факторов (повышенной температуры, по Child, Blanc and Plough, 1940) на гетерозиготу нередко выявляются признаки даже вполне рецессивной мутации. Нередко отмечалась повышенная изменчивость популяций, живущих в неблагоприятных условиях (при крайне высоких или низких температурах и т. п.).
Наконец, важнее всего то обстоятельство, что каждая новая мутация прямо или косвенно способствует выявлению имеющихся уже скрытых, а также и любых новых мутаций.
Некоторые мутации настолько изменчивы в своих выражениях, что даже употреблялись как индикаторы для анализа генетического состава линий и популяций (Тимофеев-Ресовский, Добжанский, Дубинин). У дрозофилы это касается многих мутаций, характеризующихся изменением числа щетинок или жилкования крыла, а также мутаций, выражающихся в уменьшении числа фасеток глаза. В различных генотипах они получают различное выражение, и если в нормальном генотипе они утратили свое выражение, то в любом ином они могут его восстановить. Такое восстановление наблюдалось при скрещивании лабораторных линий мутантов, утративших свое выражение (eyelles, stubbloid и др.). Реактивная способность тканей может быть вполне нормальной в довольно широких пределах между нижним и верхним порогами их раздражимости. Однако каждая мутация, сдвигающая норму реакции или меняющая концентрацию формативного раздражителя, приближает тот или иной порог нормальной реактивности. Поэтому другие мутации, которые в норме укладывались бы еще в рамки нормальной реактивности, могут у такого мутанта выйти за эти пределы. Приближение уровня фактического осуществления реакции к одному из порогов нормальной реактивности означает уменьшение «защищенности» морфогенетической реакции (уменьшение «морфогенетического запаса», по М. Камшилову). Малейший новый сдвиг в том же направлении нарушает ее нормальное течение и проявляется в виде более или менее заметного изменения строения и функций организма.
Поэтому каждая мутация, сдвигающая норму реакции в известном направлении, способствует выявлению всех мутаций, влияющих в том же направлении. Так как любая мутация обладает плейотропным выражением (хотя и добавочные выражения могут быть подавлены), то это касается не одной реакции, а в большей или меньшей мере целого ряда формообразовательных процессов. Можно смело сказать, что каждая мутация, вызывая сдвиг во времени и скорости возникновения, а следовательно — в интенсивности действия формообразовательного фактора, или, наоборот, вызывая сдвиг в реактивной способности тканей (практически это одно и то же), тем самым подготовляет почву и облегчает выявление любой старой или новой мутации, вызывающей сдвиг в том же направлении. По всей вероятности, это наиболее существенный фактор, способствующий быстрому вскрытию резервов при изменении условий существования, при котором некоторые мутации оказываются благоприятными и размножаются при содействии естественного отбора (без погашения их положительных выражений). Чем большее число мутаций при этом используется, тем большее их число приобретает явный характер.
Вскрытие резервов мутационной изменчивости всегда сопровождается значительной лабилизацией формообразования и повышением общей реактивности организма.
Процесс вскрытия уже существующих резервов имеет гораздо большее значение, чем самая интенсивная мутабильность, так как мутации, ранее накопленные в скрытом виде, проявляются сразу в значительной концентрации, обеспечивая тем самым наиболее высокую эффективность естественного отбора.
Вскрытие резервов идет, однако, с наибольшей скоростью в тех случаях, когда начинается вообще довольно свободное накопление разнообразных мутаций. Это происходит при смягчении остроты избирательной элиминации (отбора), если вид переходит в благоприятные условия существования. В этом случае увеличение изменчивости сопровождается значительным возрастанием общей численности, расширением ареала распространения и может привести к быстрой прогрессивной эволюции (ароморфоз) или дальнейшей дифференциации (алломорфоз).
С другой стороны, и при обострении общей элиминации, когда она теряет свой избирательный характер (следовательно, тоже падает острота отбора), происходит также вскрытие резервов и значительное увеличение изменчивости. Оно, однако, сопровождается в этом случае уменьшением общей численности, устранением косвенных форм соревнования и преобладанием дегенеративных изменений (катаморфоз, Шмальгаузен, 1939а).
б. Мобилизация видов. Во всех случаях вскрытие резервов означает реализацию скрытых потенций, возможность использования латентных свойств вида в процессе его дальнейшей эволюции. Эта возможность тем выше, чем больше объем резерва, т. е. чем разнообразнее эти латентные свойства. Вид приобретает известную мобильность, которая может в благоприятных условиях и при эффективном естественном отборе привести к быстрой его перестройке. Повышение изменчивости связано с увеличением эффективности естественного отбора (пропорционально о2, согласно Фишеру) и, следовательно, с возможностью быстрой эволюции.
Решающим условием для вскрытия резервов и увеличения мобильности вида в известном направлении является изменение положения организма во внешней среде. Чаще всего это связано с изменением направления естественного отбора, при котором некоторые мутации и их комбинации получают известные преимущества перед нормой (ведущая форма отбора). Происходит гомози-готизация благоприятствуемых мутаций и дальнейшее вскрытие резервов, главным образом в той их части, которая лежит в направлении действия естественного отбора. Это может произвести впечатление направленного мутирования, тем более, что к такому частичному вскрытию резервов присоединяется и выявление новых мутаций в том же направлении (так как при приближении порогового уровня данных реакций они оказываются именно в этой области «незащищенными» каким-либо морфогенетическим «запасом»). Кроме того, мы знаем, что нередко возникновение одной мутации облегчает возникновение других аллеломорфов того же ряда, вплоть до достижения наиболее устойчивого (т. е. более вероятного) состояния, которое бывает как раз наиболее удаленным от прежней нормы (например, мутация белоглазия, white, у дрозофилы). Это — обычная форма частичного вскрытия резервов и направленной мобилизации видов. Она вызывается таким же направленным изменением соотношений между организмом и внешней средой. Преобразование организма определяется ведущей формой естественного отбора и означает замену в процессе эволюции одних частных приспособлений — другими (алломорфоз, Шмальгаузен, 1939а).
Однако еще интереснее возможность общего вскрытия всех резервов и широкой мобилизации вида в любых направлениях. Это возможно в тех случаях, когда при изменении факторов среды сам отбор теряет свою строгую направленность или его влияние значительно ослабляется. Если организм теряет свою приспособленность, переходя в иную и притом разнообразную среду, в которой он встречает вместе с тем благоприятные условия для существования и размножения, то это может быть связано как раз с резким ослаблением интенсивности естественного отбора. Элиминация приобретает общий, т. е. главным образом случайный характер; в остальном она сводится лишь к гибели нежизнеспособных, т. е. негармонично построенных уклонений. Так следует представлять себе завоевание организмом новых и достаточно широких пространств в природе.
Так шло завоевание суши членистоногими и по этому же пути пошли затем первые наземные позвоночные (стегоцефалы и котилозавры). То же самое касается птерозавров и затем птиц, завоевывавших воздушную среду, водных рептилий и млекопитающих, вернувшихся в воду и нашедших здесь богатейший планктон и рыбное население океанских просторов, и многих, многих других случаев, различающихся лишь по масштабам. Образование нового водоема или острова означает освобождение нового места, которое может быть использовано с наибольшей эффективностью первыми вселенцами, если в этих местах условия делают их жизнь вообще возможной. Если условия очень благоприятны, то это ведет к бурному размножению, связанному с быстрой эволюцией, как это видно, например, по фауне Байкала с его бокоплавами, планария-ми, бычками и т. п. Однако речь идет, конечно, не только о завоевании территорий. То же самое касается и новых мест в «экономии» природы (что не всегда отделимо от понятия территории — в случае, например, воздушной или водной среды в целом), т. е. новых экологических ниш, которые не были ранее использованы или были использованы менее совершенными организмами, не могущими конкурировать с новой формой.
При ослаблении интенсивности естественного отбора, в благоприятных условиях существования, элиминация некоторых мутаций прекращается (или ослабевает) и это способствует выявлению множества «нейтральных» мутаций и их комбинаций, переходу их в гомозиготное состояние, появлению новых мутаций и более му-табильных линий. Это вскрытие резервов идет, однако, в этом случае не в одном направлении, а имеет всеобщий характер. Возрастает изменчивость популяциии во всевозможных направлениях.
По вопросу об изменчивости популяций в оптимальных условиях у экологов имеются большие разногласия. Это, несомненно, связано с неопределенностью самого понятия оптимальных условий и различием его критериев. Можно считать, что вид находится в оптимальных условиях: 1) в середине своего ареала, в котором распространение характеризуется максимальной сплошностью (биогеографический оптимум); 2) там, где население взрослых особей достигает максимальной плотности (экологический оптимум); 3) там, где смертность взрослых оказывается наименьшей и средняя длительность жизни наибольшей (физиологический оптимум); 4) там, где он наиболее обеспечен жизненными средствами и активная конкуренция сведена к минимуму, плодовитость максимальна, а элиминация имеет наиболее случайный характер. В данных оптимальных условиях все особи достаточно хорошо приспособлены, так как здесь среда не предъявляет каких-либо особых требований, кроме общей «жизнеспособности». Это последнее понимание оптимальных условий можно обозначить как биологический оптимум. В наших построениях мы оперируем с этим пониманием оптимума.
Если вид достигает в какой-либо области максимальной плотности населения (при минимальной смертности взрослых в результате малой прямой элиминации), то это означает обычно максимальную конкуренцию за жизненные средства, смертность молоди, периодическую голодовку, снижение плодовитости. Элиминация хотя и невелика, но имеет очень жесткий избирательный характер — элиминируются все уклонения от приспособленной нормы. Изменчивость в области такого оптимума может быть только минимальной (что нередко и устанавливалось).
Если мы имеем в виду наш «биологический оптимум», то для него создаются совершенно иные условия. Наше понимание оптимума не предполагает малой элиминации (и малой смертности), но предполагает, что она почти не имеет избирательного характера (малая интенсивность отбора). В этом случае плотность населения никогда не достигает предельного уровня, при котором начинается активная конкуренция за жизненные средства. Элиминация имеет вначале не косвенный, а прямой характер (потребляемость физическими факторами и хищниками подавляющей мощности). Поэтому она не приводит к снижению плодовитости, а, наоборот, статистические закономерности случайного переживания потомства поддерживают существование наиболее плодовитых линий, причем эта плодовитость при обеспеченности пищевым материалом полностью реализуется. Такой биологический оптимум возможен в двух случаях: во-первых, при свободном распространении в новой области, когда элиминация хищниками не велика, и во-вторых, даже при стационарном существовании в определенных условиях, когда прямая потребляемость очень велика, но не имеет избирательного характера.
Первый случай ограничивается во времени лишь периодом продолжающейся экспансии. Однако он наиболее интересен, так как сопровождается наиболее бурной мобилизацией и быстрой эволюцией по пути общего повышения организации (ароморфоз) или частного приспособления (алломорфоз).
Резкое ослабление остроты отбора (за исключением отбора на «общую жизнеспособность») ведет к быстрому вскрытию резервов изменчивости и повышению общей мутабильности. Изменчивость популяции достигает своего максимума в период ее расселения и увеличения численности на новых местах. Однако вскоре начинается либо размножение имевшихся уже хищников или паразитов, либо приспособление новых форм к хищничеству или паразитизму на новом иммигранте. Тогда элиминация приобретает избирательный характер и начинается очень быстрая (при большой эффективности отбора) эволюция в различных направлениях выработки мер защиты, частных приспособлений и т. д. Если же в завоеванной новой области действительно нет хищников, уничтожающих новую форму (как было, например, при первом завоевании суши членистоногими, а затем — позвоночными и вообще при всяком вторжении в еще необитаемые области новых водоемов или новых островов), то прогрессивное увеличение плотности населения ведет к развитию конкуренции, т. е. хотя и косвенной, но строго избирательной элиминации. В качестве примера можно сослаться на богатейшее видообразование рыб рода Haplochromis в озере Виктория, где не было активных хищников (Worthington, 1940), а также на видообразование бычков в оз. Байкал. Эффективный в этом случае отбор ведет к максимальной дифференциации при расхождении признаков и дальнейшей специализации, связанной со строжайшим распределением ролей (адаптивная радиация новых форм).
Во втором случае, когда прямая истребляемость велика и не допускает большого увеличения численности, но вместе с тем и не имеет избирательного характера (ввиду мощности стихийных сил неорганической или органической природы — климата или хищников подавляющей силы), повышение изменчивости приобретает длительный характер. Большая цифра гибели компенсируется высокой плодовитостью. Свободное накопление мутаций ведет к дегенеративным явлениям, к измельчанию, общему недоразвитию или преждевременному созреванию, в которых иногда и создается выход из создавшегося трудного положения (скрытая жизнь измельчавших форм). Это — путь катаморфоза, который может привести, при усиливающемся истреблении, и к «дегенеративному» вымиранию (Шмальгаузен, 1939а).
Ясно, что чем выше объем мобилизационного резерва, т. е. чем выше развит регуляторный аппарат организма, допускающий в широком масштабе скрытое мутирование и накопление нейтрализованных мутаций, тем выше потенциальная мобильность организма. Соответственно наиболее мобильны позвоночные животные, которые и достигали максимально высоких темпов эволюции при благоприятных для этого условиях.
Скорость вскрытия резервов и возможность их мобилизации очень хорошо видны на процессах одомашнения животных и введения в культуру различных используемых человеком растений. При условии ухода и охраны со стороны человека и главное подкармливания, у домашних животных потеряли свое значение почти все элиминирующие факторы. Естественный отбор если и не прекратился, то почти утратил свое влияние; остался лишь отбор на общую «жизнеспособность» в создаваемых человеком условиях. Поэтому должно было идти быстрое вскрытие резервов, которое затем и привело при участии искусственного отбора к быстрому породообразованию. В нем участвовали и новые мутации, возникавшие в особенности в благоприятствуемом направлении. Предел быстрого начального формообразования был, очевидно, продиктован объемом накопленного ранее резерва. Предел дальнейшей селекции диктуется физиологическими условиями «жизнеспособности» организма.
Ускорению мобилизации резервов и их обогащению способствовала также гибридизация, издавна применявшаяся человеком и приводившая к нарушению внутриклеточного «генного баланса» и к некоторому расшатыванию прочих регуляторных механизмов.
Мало специализированные исходные формы, жившие в благоприятных условиях и обладавшие большим запасом изменчивости, дали начало очень большому разнообразию пород — домашняя курица, собака, овца. Специализированные формы, жившие в более жестких условиях борьбы за существование и обладавшие соответственно меньшим запасом изменчивости, оказались менее мобильными и не дали такого пышного породообразования — домашняя утка и гусь (перелетные птицы с периодически более жестким отбором), кошка (специализированный хищник с острой конкуренцией; гибридизация, очевидно, не происходила), лошадь (специализированное травоядное с острой зимней элиминацией).
Из культивируемых растений скрытый запас изменчивости должен был быть выше у полиплоидов. Большое число сортов пшениц связано, очевидно, с их полиплоидностью. То же самое относится и к наиболее распространенному декоративному растению — культурным сортам роз. Впрочем, у растений общая картина сложнее, так как к этому присоединялись не только более свободная гибридизация и усиленное мутирование в условиях, создаваемых искусственными средствами культивирования, но также и различные другие факторы, в том числе соматические мутации, используемые методами вегетативного размножения и прививок. Все это способствовало еще более полному выявлению латентных свойств, чем это было возможно у животных.
Во всяком случае, и породообразование домашних животных и сортообразование культивируемых растений произошло с такой исключительной скоростью, очевидно, главным образом за счет накопленного ранее резерва изменчивости. Дальнейшая строго направленная селекция идет уже медленнее, хотя возможность выявления более мутабильных линий, а также гибридизации и влияния внешних воздействий позволяют и здесь рассчитывать на гораздо большие скорости и не ставит в этом смысле никаких ограничений (за исключением физиологически допустимых пределов).
в. Иммобилизация и специализация видов. Когда распространившаяся новая форма осела на новых местах или в новых экологических нишах и достигла известной плотности населения, начинает иногда сказываться постоянная или временная, периодическая или случайная нехватка жизненных средств. Тогда борьба за существование обостряется и в особенности возрастает активное соревнование особей за жизненные средства. В этом случае элиминация приобретает все более индивидуальный характер. Естественный отбор на общую жизнеспособность (при наиболее обеспеченных условиях существования) переходит в отбор на наибольшую организованность (выносливую конституцию) и приспособленность. Чем острее конкуренция, тем далее идет экологическое расхождение, тем более частный характер приобретает приспособленность организма, переходящая мало-помалу во все более узкую специализацию (теломорфоз).
Ведущая форма отбора теряет свое значение у максимально приспособленного организма и уступает место стабилизирующей форме. Влияние последней несколько различно в различных условиях борьбы за существование. Если лабильный организм не слишком специализирован и живет в разнообразной среде или если он даже специализирован, но живет в условиях постоянных и случайных изменений (непериодических колебаний) в факторах внешней среды, то стабилизирующий отбор (элиминация вредных мутаций и неблагоприятных модификационных уклонений от приспособленной нормы) ведет именно к стабилизации, т. е. к максимальной устойчивости формообразования в условиях изменчивой среды. Если же специализированный организм живет в однообразных и относительно постоянных условиях среды, то стабилизирующий отбор (т. е. элиминация всех — в данных условиях именно наследственных — уклонений от нормы) ведет к иммобилизации, т. е. к утере эволюционной пластичности. Иммобилизация есть результат острой элиминации всех мутаций, выражающихся в уклонениях от нормальной организации и ее нормальных функций. Она препятствует накоплению новых резервов изменчивости. Элиминируются, однако, именно проявляющиеся мутации, т. е. мутации, имеющие морфофизиологическое выражение. Поэтому иммобилизация достигает различного уровня у организмов, отличающихся по степени развития своего регуляторного аппарата.
Наиболее значительной должна быть иммобилизация гаплоидного организма, так как при отсутствии важнейших внутриклеточных регуляторных механизмов, связанных с диплоидно-стью, почти любая мутация должна иметь свое выражение и поэтому подлежит элиминации. Таким образом, гаплоидный организм не может иметь значительного скрытого мобилизационного резерва.
Редукция гаплоидного поколения у растений связана, вероятно, именно с его иммобильностью, т. е. эволюционным консерватизмом.
Так как мутации получают, во всяком случае, более значительное выражение при их переходе в гомозиготное состояние, то и гомозиготизация ведет к усиленной элиминации наследственных уклонений, и следовательно, к иммобилизации специализированных организмов. Все, что связано с родственным размножением, ведет поэтому к иммобилизации видов и утере эволюционной пластичности. Поэтому все формы более полной изоляции небольших популяций означают вместе с тем их иммобилизацию. В особенности, конечно, в тех случаях, когда мы имеем дело со специализированным организмом, живущим в весьма определенной и относительно константной среде и обладающим притом слабо развитой регуляторной системой (например, у животных с мозаичным развитием).
Консерватизм очень многих беспозвоночных связан с их специализацией и с жизнью в весьма однородной и постоянной среде. Это касается в особенности морских животных, живущих в защищенной обстановке, или глубоководных, а также малоподвижных и сидячих форм с малой потребляемостью (т. е. с преобладанием косвенной элиминации). Сидячие кишечнополостные, в том числе кораллы, многие двустворчатые и брюхоногие моллюски, частью и иглокожие, сидячие кольчецы, а в особенности брахиоподы отличаются удивительным консерватизмом на протяжении долгих геологических периодов. Этому способствовал и мозаичный характер развития высших червей, моллюсков и брахиопод. Исключение составляют головоногие моллюски — подвижные и активные хищники, обладающие хорошо развитым регуляторным аппаратом. Соответственно, они оказались в общем достаточно мобильными. Из хордовых животных консервативными оказались оболочники — специализированные организмы с мозаичным развитием, а наиболее мобильными — позвоночные, особенно высшие позвоночные, гораздо более универсальные по своей организации и обладающие наиболее развитой системой морфогенетических корреляций регу-ляторного характера и соответственно, очевидно, значительным запасом наследственной изменчивости,
Шпет и Иванько (1940) сделали попытку сопоставить изменчивость представителей консервативных («древних») групп насекомых (Odonata, Blattoidea, Ephemeroptera) и изменчивость представителей прогрессивных групп (Diptera, Hymenoptera, Lepidop-tera). Получились биометрически ясные различия в пользу последних.
В практике разведения домашних животных и культивируемых растений мы имеем дело также со специализацией и иммобилизацией пород и сортов. Благоприятные условия размножения животных и культивируемых растений при неограниченных жизненных средствах и защите со стороны человека вели вначале ко вскрытию резервов изменчивости и их мобилизации. Это привело при применении искусственного отбора (в его «ведущей» форме) к быстрому породо- и сортообразованию. Однако дальнейшая, более строгая, селекция шла по пути более узкой и притом неограниченно далеко идущей «специализации» требований (отбор мясных, шерстных и т. д. пород), что привело к выработке стандартов и ко все более интенсивной браковке уклонений от этих стандартов. Это можно рассматривать как стабилизирующую форму искусственного отбора.
Однако искусственная стабилизация ведется теперь и в специально выработанных (пусть даже оптимальных) стандартных условиях развития и содержания. Следовательно, стабилизирующий отбор происходит в относительно весьма постоянных условиях внешней среды. Поэтому почти все уклонения имеют наследственный характер. Элиминация наследственных уклонений означает тогда именно иммобилизацию, т. е. утрату пластичности. Той же иммобилизации содействует, конечно, и внутрипородное родственное разведение. Впрочем, у домашних птиц и млекопитающих, обладающих высоко развитой системой морфогенетических регуляций, иммобилизация не может быть полной, так как скрытое мутирование ускользает от контроля человека.
То же самое происходит и с лабораторными и любыми экспериментальными культурами. Если они вначале ставятся в оптимальные условия существования и размножения, то имеет место быстрое вскрытие резервов — увеличение изменчивости. В дальнейшем происходит выделение известных линий и их стабилизация, а в условиях родственного разведения в выработанных стандартных условиях — их иммобилизация. В особенности это касается культур животных с мозаичным развитием и слабо развитой системой регуляций, как, например, дрозофилы.
Свеже поставленные культуры дрозофилы, взятой из природных популяций, показывают всегда вскрытие резервов (более значительных, если это большая популяция, менее значительных, если это небольшие изолированные популяции, и в особенности малых, если популяции жили в постоянных, ограниченных и суровых условиях острой элиминации). Однако лабораторная линия «Флорида», прошедшая уже в лабораториях огромное число поколений, из которых систематически элиминировалась каждая замеченная мутация, прошла таким образом путь жесткой стабилизации.
Так как эти культуры ведутся в стандартных условиях постоянной температуры (25°), постоянной влажности и стандартной кормовой смеси, то «стабилизирующий» искусственный отбор привел затем к неизбежной «иммобилизации». Лабораторная линия «Флорида» естественно теряет свою пластичность. По Р. Л. Берг, она так же мало мутабильна, как и вполне изолированные небольшие природные популяции дрозофилы. По мере дальнейшей культуры в стандартных условиях можно ожидать лишь усиливающегося снижения мутабильности.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 2254; Нарушение авторских прав?; Мы поможем в написании вашей работы!