КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Контроль и регуляция в эволюции 1 страница
|
|
|
|
А. ЭВОЛЮЦИЯ КАК РЕГУЛИРУЕМЫЙ ПРОЦЕСС
Эволюция, т. е. процесс исторического развития организмов, протекает закономерно. Основные закономерности этого процесса были вскрыты еще Ч. Дарвином. Они выражаются в непрерывном приспособлении организмов во всех своих функциях и всех чертах своей организации к той среде, в которой они обитают и именно к тем факторам среды, с которыми они сталкиваются в своей жизнедеятельности. Это приспособление достигает иногда удивительного совершенства и исключительной тонкости и вместе с тем оно всегда совпадает именно с теми условиями, в которых протекает жизненный цикл данного организма. Если в течение жизненного цикла происходит закономерная смена среды обитания (например, переход от водной жизни к наземной, как у амфибий и у некоторых насекомых, или от скрытой жизни в земле или в растительных тканях к открытой жизни, как у многих жуков), то это всегда сопровождается соответствующей перестройкой организации. Если у организма имеются ненужные ему органы и приспособления, это указывает на исторический выход из другой среды и всегда ведет к постепенной редукции этих органов и приспособлений (рудиментарные органы). Такое точное приспособление организма к внешней среде не было бы возможно, если бы не существовало механизма, контролирующего и регулирующего исторический ход этого приспособления. Этот механизм управляет, очевидно, процессом эволюции организмов в связи с факторами внешней среды. Должна быть органическая связь между организмом и средой не только в его жизни (что и без того ясно), но и в ходе их исторических преобразований. Такая связь была найдена Ч. Дарвином в виде борьбы всех живых существ за свое существование в конкретной жизненной обстановке. Борьба за существование в условиях конкуренции в овладении жизненными средствами ведет к переживанию наиболее приспособленных особей любого вида организмов и их преимущественному размножению. В этом и состоит естественный отбор как основная часть регулирующего механизма эволюции. Теория естественного отбора полностью оправдала себя в течение дальнейшего развития биологических знаний. Она, однако, не дает полного объяснения всего эволюционного процесса.
|
|
|
Естественный отбор — это регулирующий механизм эволюции, но не ее исходная материальная основа. Ч. Дарвин указал и на эту основу — изменчивость и наследственность. Однако закономерности изменчивости и наследственности, несмотря на огромный эмпирический материал, собранный Дарвином, оставались в то время еще совершенно неизвестными. Только текущее столетие пролило свет на эти проблемы. Благодаря блестяшчм экспериментальным исследованиям, берущим свое начало от классических опытов Менделя, мы можем в настоящее время сказать, что в основном механизм изменчивости и наследственности уже вскрыт.
Таким образом, в настоящее время хорошо изучена материальная база эволюционного процесса и известен механизм его регуляции. Это — начальное и конечное звенья в элементарном цикле эволюционных преобразований. Роль этих звеньев достаточно ясна и легко поддается количественному учету. Для этого были введены понятия мутационного «давления» (т. е. скорости или частоты возникновения наследственных изменений) и «давления» естественного отбора (т. е. его эффективности в известных условиях). Эти понятия легли в основу генетической теории естественного отбора (Fisher, 1930). Вместе с экспериментальными исследованиями популяционноп генетики они положили начало точному анализу факторов эволюции. Несмотря на большое значение уже достигнутых результатов, такие исследования не вскрывают всего механизма эволюции и не дают полного объяснения его закономерностей.
|
|
|
При таком подходе в тени остается индивидуальное развитие организмов, ведущее к реализации фенотипа. Так как именно фенотипы являются активными носителями жизни и объектами естественного отбора, то ход индивидуального развития не может не иметь значения эволюции. И, наконец, самое главное, в генетической теории естественного отбора не видно организма как такового с его активной борьбой за свою жизнь. Понятие борьбы за существование, лежащее в основе теории Дарвина, совершенно выпало. Естественный отбор выступает как внешний фактор, а сам организм — как пассивный объект, с которым оперирует естественный отбор. Это не является верным отражением действительных соотношений. В оправдание установившейся в генетике точки зрения следует сказать, что дарвиновское понятие борьбы за существование сформулировано столь расплывчато, что с ним действительно трудно оперировать. Это видели многие критики и особенно К. А. Тимирязев, который предложил вообще заменить его понятием элиминация. Последняя поддается точному учету. Однако элиминация является лишь обратной стороной естественного отбора и в таком виде остается внешним фактором по отношению к организму.
Нужны, очевидно, новые понятия, во-первых, отражающие активную роль организма не только в своих жизненных проявлениях, но и в эволюции и, во-вторых, хорошо ограниченные и доступные точному исследованию. Возможно, что такие понятия удастся сформулировать, если подойти к эволюции с новыми представлениями об устройстве регулируемых механизмов вообще, т. е. с точки зрения кибернетики (Ashby, 1956) (включающей теорию информации). Как уже пояснено, любой автоматически регулируемый механизм составляется из двух частей — регулируемого объекта и регулятора, которые соединены между собой линиями связи, передающими управляющие сигналы от регулятора и обратную информацию о состоянии объекта к регулятору.
В процессе эволюции органического мира управляемым объектом является вид в целом. Однако вид организмов обычно разбит на подчиненные единицы, ведущие до известной степени самостоятельное существование и обнаруживающие нередко явные признаки приспособления к местным условиям. Наименьшей из таких единиц является популяция особей. Поэтому, рассматривая лишь элементарные основы эволюции, мы должны исходить из популяции как наименьшей эволюирующей единицы. Направление эволюционного процесса определяется, очевидно, внешней средой. На это ясно указывает точная приспособленность организмов к условиям их существования и при том не только к климатическим, эдафическим и другим факторам неорганического окружения, но и к тем живым существам, с которыми они сталкиваются в этой среде и с которыми они связаны теснейшими взаимоотношениями (и прежде всего пищевыми связями). С другой стороны, нельзя преуменьшать и роли внутренних факторов. На их значение указывает точная слаженность организации, взаимоприспособленность отдельных частей, органов и тканей, согласованность их функций. Наконец, и в выборе местообитания, и в столкновениях с другими организмами, и в пищевых взаимоотношениях активную роль играют и сами особи данной популяции.
|
|
|
Таким образом ясно, что регулирующий механизм кроется в некоторых формах взаимодействия между организмом и внешней средой. Этот механизм и был вскрыт Ч. Дарвином. Его действие проявляется в различных формах борьбы за существование, ведущей через дифференциальное переживание и размножение к естественному отбору наиболее приспособленных особей. Борьба за существование проявляется в теснейшем взаимодействии организма и среды (включающей все другие организмы, совместно с ним обитающие и образующие вместе с ними единое «сообщество» — биоценоз). Так как мы не можем отрицать значение факторов неорганической среды, в которой живет и размножается данный организм, мы будем говорить о биогеоценозе (Сукачев, 1945) как о регулирующем аппарате эволюции. Любая популяция особей определенного вида, входящая в состав известного биогеоценоза, является его органической, необходимой составной частью. Таким образом, в данном случае обнаруживается самая интимная связь между управляемым объектом — популяцией и регулятором — биогеоценозом. Такая тесная связь наблюдается нередко и в конструируемых нами механизмах, где управляемый объект является как бы частью регулятора или, наоборот, регулятор входит как часть в состав управляемой машины. Существование тесной взаимосвязи и является условием, допускающим возможность контроля и управления эволюционным процессом. Если биогеоценоз в целом играет роль регулятора эволюционного процесса, то он обязательно должен быть обеспечен «информацией» о состоянии популяции (по линии «обратной» связи), должен включать в себя специфический механизм преобразования этой информации в управляющие сигналы и средства передачи последних на популяцию. Таким образом, кроме механизма преобразования требуется наличие каналов связи для передачи информации в двух направлениях — от популяции к биогеоценозу и от биогеоценоза к популяции. Так как изменение популяции, как элементарный эволюционный процесс, всегда сопровождается наследственным изменением ее особей, то управляющие сигналы от биогеоценоза к популяции должны каким-то образом включать возможность изменения ее наследственной структуры. Последнее может произойти только в процессе преобразования информации в самом биогеоценозе (т. е. в «регуляторе»). Так как первичные эволюционные изменения возможны только в популяции (или в поколениях особей, но не в отдельных особях), то наиболее простым изменением является хотя бы небольшое изменение в генетическом составе популяции, т. е. в соотношении числа особей с разной наследственной характеристикой (генотипов). Информация о таких изменениях популяции может быть сообщена через наследственный аппарат ее особей и передана особям следующего поколения при посредстве половых клеток или иных средств воспроизведения. Такой аппарат действительно имеется и несомненно он полностью обеспечивает надежную связь популяции с регулирующим механизмом биогеоценоза и дальнейшую передачу информации от одного поколения особей к следующему. Имеются и средства передачи обратной информации от популяции к биогеоценозу.
|
|
|
Популяция, несомненно, активно воздействует на биогеоценоз, хотя бы через потребление пищевых материалов и накопление продуктов своей жизнедеятельности. В известных условиях популяция может внести значительные изменения в строение биогеоценоза. Таким образом, имеются и каналы обратной связи.
Однако нет прямой связи между наследственной информацией по первому каналу (от биогеоценоза) и обратной информацией по второму каналу (от популяции к биогеоценозу). Здесь непосредственная связь как будто прерывается, так как обе линии связи находятся на разных уровнях. Наследственная информация передается на внутриклеточном (молекулярном) уровне организации, а обратная информация только на уровне организации целой особи.
Переход от одной линии связи к другой совершается здесь (как обычно и в технике) через посредство довольно сложного механизма преобразования. Наследственная информация преобразуется в процессах индивидуального развития в средства передачи обратной информации, именно — в фенотип особи, являющейся реальным носителем жизни и активным участником наступления на жизненные ресурсы биогеоценоза («борьбы за существование»). В биогеоценозе через естественный отбор и процессы размножения происходит новое преобразование этой информации в наследственную с переходом от уровня организации особи (в фенотипах) на уровень организации клетки (половые клетки, зиготы). Этим и замыкается полный круг преобразований в элементарном цикле эволюционного процесса (табл. 3).

Таким образом, основные требования, предъявляемые к конструкции автоматически регулируемого устройства, в данном случае оказываются выполненными, и мы можем попытаться осветить более подробно работу отдельных звеньев этого механизма. Это необходимо сделать для того, чтобы представить себе яснее, может ли новая точка зрения внести что-либо новое в понимание эволюционного процесса, выдвигает ли она новые вопросы, освещает ли она наиболее ответственные звенья этого процесса. Не позволит ли она сформулировать основные понятия таким образом, чтобы внести полную ясность и четкость в понимание механизма эволюции и сделать все его звенья доступными для точного количественного анализа.
Б. НАСЛЕДСТВЕННАЯ ИНФОРМАЦИЯ И СРЕДСТВА ЕЕ ПЕРЕДАЧИ
Наследственная информация передается от материнских особей к дочерним особям, т. е. от предыдущей генерации к последующей генерации той же популяции. В предшествующем изложении мы говорили также о передаче информации от биогеоценоза к популяции. Нет ли здесь неувязки? Нет, неувязки здесь нет, но нужно учесть, что наследственная информация передается от предыдущей генерации к последующей только после ее преобразования в биогеоценозе. Схема популяции входит в состав биогеоценоза в качестве его неотъемлемой части. Механизм контроля и преобразования информации заключается в самом биогеоценозе, именно во взаимодействии особей данной популяции с другими членами и факторами биогеоценоза. Преобразование совершается через естественный отбор, т. е. при преимущественном размножении одних особей с их наследственными механизмами и элиминации других. Таким образом, под влиянием естественного отбора в самом биогеоценозе происходит изменение состава популяции, меняется ее наследственная характеристика. Во всей популяции в целом создаются новые соотношения по меньшей мере в численности различных генотипов. Механизм наследственной передачи лежит, следовательно, в отдельных особях, а изменение в биогеоценозе касается не отдельных особей, а всей популяции в целом. Информация об этих изменениях передается все же через посредство размножения апробированных особей, потомство которых вливается в ту же популяцию. Таким образом меняется популяция, но информация об этом изменении (происшедшем в биогеоценозе) передается через размножение особей.
Средством передачи наследственной информации являются либо половые клетки, либо другие пропагационные клетки, образующие зачатки нового организма при вегетативном размножении. Во всяком случае такая передача осуществляется только через посредство клеток. Механизм этой передачи хорошо известен, он совпадает с механизмом клеточных делений, при котором полностью обеспечивается равномерное распределение основных ядерных субстанций в виде определенного набора хромосом. Каждая клетка получает полный набор этих хромосом. Изучение строения последних с помощью двух принципиально различных методов — гибридологического и цитологического — показало их сложный состав из большого числа наследственных единиц, располагающихся в ряд по всей длине хромосомы. Гибридологической единицей наследственности является ген. Изучение химического состава хромосом показало, что важнейшей их составной частью является дезоксирибонуклеиновая кислота (ДНК). Молекула последней имеет вид очень длинной парной цепочки, состоящей из чередующихся фосфатных и сахарных групп, связанных парами пурино-вых и пиридиновых оснований, образующих как бы перекладины веревочной лестницы (Watson, 1953). Строение этих гигантских молекул обеспечивает как высокую индивидуальность, так и значительную их стойкость. Оно дает также возможность продольного расщепления молекулы с последующим восстановлением каждого партнера. Очевидно, такое расщепление и восстановление и лежит в основе расщепленияхромосом и их генов при клеточных делениях. Во всяком случае это гарантирует возможность точной передачи наследственной информации от клетки к клетке.
Вся полнота информации о наследственных свойствах зиготы и развивающейся из нее особи обеспечивается определенной последовательностью и положением двух возможных пар оснований (аденин с тимином или гуанин с цитозином) в каждой молекуле. При большом числе звеньев, исчисляемом тысячами, комбинированное расположение четырех качественно различных знаков (оснований) гарантирует возможность записи наследственных структур любой сложности в виде условного «кода». Наследственная информация передается, следовательно, в закодированном виде и при том вся единовременно в пространственной связи, а не во временной последовательности. При расшифровке этого кода в процессах индивидуального развития информация развертывается также в пространстве, хотя ее преобразование совершается несомненно во временной последовательности. Химической единицей нашего кода является, как видно, пара оснований, которая может принять форму лишь четырех знаков (аденин с тимином, гуанин с цитозином, или в противоположном положении — тимин с аденином и цитозин с гуанином). Чередование этих знаков может обеспечить передачу любой информации (как телеграф Морзе с таким же числом знаков). Гибридологической единицей наследственности является, однако, более сложное образование — ген как выражение системных связей в хромосомах (об этом см. дальше в разделе Ж).
В. ПРЕОБРАЗОВАНИЕ ИНФОРМАЦИИ В ИНДИВИДУАЛЬНОМ РАЗВИТИИ
Преобразование информации представляет всегда закономерный процесс, полностью определяемый устройством той системы, в которой совершается это преобразование. Наследственная информация, заключающаяся в зиготе или в другой пропагационной клетке, преобразуется затем в процессах индивидуального развития. Характер преобразования определяется организацией зиготы и поэтапно развивающейся из нее особи. Это преобразование представляет собой очень сложный процесс (полное рассмотрение которого выходит за пределы поставленной нами задачи). Он строится на биохимической основе внутриклеточного обмена, регулируемого ядерными субстанциями хромосом. Однако в эти процессы, естественно, вовлекаются и плазменные субстанции, взаимодействующие с ядерными продуктами в процессах этого обмена (табл. 4). Первоначальные дифференцировки зиготы и дробящегося яйца определяются именно организацией плазмы, детерминированной еще в яичнике материнского организма. Дальнейшая дифференцировка идет иногда при заметном участии внешних факторов, но определяется все же взаимодействием частей развивающегося организма. Такие взаимодействия частей осуществляются в «индукционных», «формообразовательных» и «корреляционных» системах, которые имеют более или менее ярко выраженный регуляторный характер (гомеостат). Внешние факторы не имеют специфического формообразовательного значения. Они оказываются лишь в роли освобождающих причин, определяющих время и иногда место наступления известной реакции, и в лучшем случае служат для включения «стрелочного» механизма, направляющего развитие по одному или другому из существующих уже, т. е. полностью детерминированных, путей («каналов») развития. На более поздних стадиях развития регуляция формообразования переходит к эндокринной системе и к самой функции отдельных органов, связанных через посредство кровеносной и, в особенности, — нервной системы. В функциональных зависимостях вновь ясно выражается связь с внешней средой и влияние факторов среды на формообразование. Однако это влияние всегда является косвенным и не определяет специфики морфологических процессов. Таким образом, если некоторые функции контроля и выполняются внешней средой, то все же специфические ответы на факторы этой среды определяются унаследованными нормами реакции данного организма. Только поэтому фенотип особи и является полным и точным (для данных условий среды) выражением генотипа. Регуляторные функции выполняются всегда унаследованным механизмом развития самого организма. Поэтому всякое развитие организма (особи) есть по меньшей мере авторегуляция. В высших формах индивидуального развития зависимость от факторов внешней среды теряет в еще большей мере свое формообразовательное значение и авторегуляция переходит в более или менее ярко выраженное автономное развитие. Как и все формы преобразования информации, индивидуальное развитие определяется в своей специфике только организацией самого преобразующего механизма. Во всех случаях формообразования оно индуцируется очень незначительным влиянием, небольшой разницей в обмене, незначительным количеством специфических продуктов индуктора или эндокринной железы, ничтожным поступлением энергии из внешней среды и реализуется в полной мере при малейшем превышении порогового уровня реактивности материала. Реакция ни в какой мере не пропорциональна формообразовательному воздействию. Она осуществляется на известном его уровне сразу и в полном масштабе, определяемом унаследованной «нормой». Обычно она выражается в клеточных делениях, перемещениях и дифференцировке больших клеточных масс. Реакция имеет по существу «взрывной» характер. Она подготовлена заранее в виде известного запаса энергии внутри реагирующего материала. Такой характер индивидуального развития, т. е. преобразования наследственной информации, напоминает обычный метод преобразования информации с использованием специфических усилителей в регулирующих механизмах автоматического управления.
Наследственный материал хромосом является средством передачи унаследованной структуры (т. е. передачи закодированной информации по первому каналу связи) от материнского организма через половую клетку и зиготу ко всем клеткам дочернего организма. Хромосомы являются, однако, не только средством механической передачи химических веществ определенной унаследованной структуры от клетки к клетке. Хромосомы являются прежде всего активными регуляторами внутриклеточного обмена веществ и контролируют через это все процессы индивидуального развития. Наследственная информация реализуется через эти процессы в виде развивающейся особи — фенотипа. Фенотип является внешним выражением генотипа и вместе с тем активным руководителем внеклеточного обмена веществ между организмом и внешней средой. Он контролирует всю его жизнедеятельность. И вместе с тем фенотип является средством обратной информации от организма к внешней среде (биогеоценозу). Эта передача производится совсем иными средствами, чем передача наследственной информации от материнского организма к дочернему. И все же она находится в органической связи с этой передачей — фенотип вполне закономерно отображает генотип, но только в совершенно иной форме. В индивидуальном развитии происходит преобразование скрытой, наследственной информации (записанной в виде условного кода в хромосомах) в явную информацию об унаследованных нормах реакции. Это преобразование происходит во взаимодействии с факторами внешней среды, которые являются необходимыми условиями и, иногда, освобождающими факторами этого преобразования. Вся его качественная специфика в данных условиях определяется, однако, унаследованными нормами реакций, т. е. генотипом. Поэтому фенотип является вполне реальным выразителем генотипа в известных условиях развития. Фенотип есть преобразованный генотип и если последний закодирован в тончайшей структуре хромосом и управляет внутриклеточным обменом (т. е. жизнедеятельностью клетки), то первый обнаруживается во всех признаках целого организма (особи) и проявляется в специфических формах его жизнедеятельности. Наследственная структура хромосом является средством межклеточной связи внутри организма (от зиготы к первичной половой клетке через многочисленные клеточные деления) и регулятором внутриклеточного обмена веществ. Фенотип является средством связи между особью и внешней средой и регулятором обмена веществ между особью и этой средой. В обоих случаях эти структуры ответственны за основные жизненные проявления, но только хромосомы — в масштабе клетки, а фенотип — в масштабе целой особи. Фенотип также «записан» условными символами, как и генотип, но только это совершенно иные символы, так как они несут хотя и сходную функцию, но на различных уровнях организации в разных условиях среды.
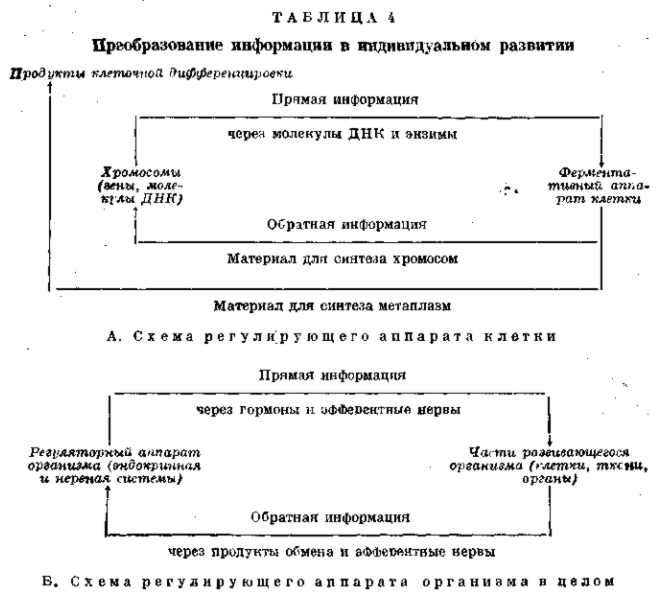
Наследственная информация, передающаяся на молекулярном уровне организации и реализующаяся в процессах внутриклеточного обмена, преобразуется в обратную информацию, передаваемую на уровне организации особи и реализующуюся в процессах жизнедеятельности, т. е. обмене веществ целой особи. Весь смысл индивидуального развития состоит в преобразовании наследственной информации в систему жизненных связей организма с внешней средой. Эти связи устанавливаются посредством признаков фенотипа, которые одновременно являются и средствами, позволяющими осуществить контроль этой организации со стороны факторов биогеоценоза. Поэтому индивидуальное развитие организма, связанное с реализацией (в известных условиях) наследственной структуры в фенотипе развивающейся, а затем и зрелой особи, представляет типичный пример преобразования информации с переходом от одного кода к другому.
Г. ПЕРЕДАЧА ОБРАТНОЙ ИНФОРМАЦИИ ОТ ПОПУЛЯЦИИ К БИОГЕОЦЕНОЗУ
Средством передачи обратной информации является фенотип и именно специфические формы активности каждой особи, как разнообразные выражения ее жизнедеятельности в определенной обстановке— в биогеоценозе.
Основным проявлением активности является добывание жизненных средств и прежде всего потребление пищевых материалов (пищевые материалы используются на развитие, рост, на поддержание жизни и на затраты энергии при всех проявлениях жизнедеятельности). Вторым проявлением активности является деятельность, связанная с размножением — половая активность и воспроизведение потомства. Третьим проявлением жизненной активности является деятельность, связанная с защитой своей жизни и жизни потомства. Обратная информация передается, таким образом, через весь фенотип в целом. Это не значит, что эта информация неразложима. Как геном при всей своей целостности разложим на единицы наследственной информации — гены и даже еще далее — на простейшие химические единицы (пары оснований), так и целостный фенотип разложим на многочисленные компоненты, являющиеся также своего рода символами, отображающими наследственные свойства особи. Связь между этими символами или признаками является весьма тесной. И все же некоторые признаки могут быть разделены и перекомбинированы или изменены в результате соответствующих изменений в наследственных единицах — генах. Это касается прежде всего таких поверхностных признаков, как окраска, рисунок, структура покровов, форма и окраска волос и многие другие. Отдельные признаки внутренней организации также могут быть изменены и это приводит к изменениям в обмене, в функциях отдельных органов, в поведении. Чаще всего, однако, такие изменения затрагивают целые взаимосвязанные системы, регулирующие основные жизненные проявления, и тогда их результат сказывается в изменении общей выносливости организма по отношению к внешним факторам (климату), общей активности, жизнедеятельности, плодовитости.
Взаимосвязанность большинства признаков фенотипа является условием жизнеспособности, устойчивости и активности особи и вместе с тем условием, обеспечивающим высокие качества передачи информации. В каждой особи признаки фенотипа обнаруживают высокую степень «сцепления». Они настолько связаны, что образуют в сущности одну целостную систему. Каждая особь глубоко индивидуальна и является надежным носителем высококачественной информации. Последняя имеет и в этом случае (как и в геноме) «картинный» характер, она развертывается не во временной последовательности, а в пространстве. Хотя она передается и во времени, но при этом различные ее комплексы выявляются все же сразу. Во времени меняется и фенотип и, следовательно, сама информация (возрастные изменения), но и в этом случае меняется одновременно вся «картина». Обратная информация от популяции передается через посредство не одной особи, а всех особей данной популяции. Особь является лишь самостоятельной единицей этой информации. Поэтому вся информация оказывается гораздо более сложной. При максимальном разнообразии особей, когда каждый фенотип (как и генотип) неповторим, количество информации является максимальным. Во вполне однородной популяции количество информации падает до нуля и регуляция ее состава оказывается невозможной.
Особи являются носителями равновероятных сообщений, если фенотипы равномерно распределены в популяции. Они могут, однако, группироваться по некоторым признакам фенотипа в подчиненные единицы разной численности (полиморфизм). Такие группы получают свою качественную характеристику, могут получить положительную оценку в известных условиях и дать начало новой самостоятельной популяции и новому таксономическому подразделению. Особи одной популяции являются, как правило, изолированными носителями сходной информации. Этим гарантируется повторность и, следовательно, надежность информации по наиболее существенным признакам организации. Одновременно по второстепенным признакам этой повторности нет, так как в этом отношении проявляются большие различия между особями. Многообразие особей при их изолированности делает состав популяции весьма подвижным — в процессе эволюции он легко меняется (через контроль и процесс преобразования информации в биогеоценозе).
Однако не всегда особи одной популяции являются вполне изолированными единицами. В некоторых случаях между особями данной популяции или ее части устанавливается более тесная связь. Выделяются временные группы особей, называемые стадами, стаями, косяками или, при более тесной и постоянной связи по местообитанию — колониями. Такое объединение особей ведет к улучшению качества информации по меньшей мере в результате увеличения помехоустойчивости (см. далее). Попарное объединение особей разного пола ведет к более надежной передаче информации следующему поколению (по первому каналу), но связано и с некоторым повышением помехоустойчивости в передаче обратной информации.
Передача обратной информации от популяции к биогеоценозу осуществляется, как сказано, через посредство отдельных особей и выражается в различных формах жизненной активности. Прежде всего эта активность проявляется в добывании жизненных средств. Она строго специфична, так как особи одного вида потребляют совершенно определенные неорганические и органические вещества. Она до некоторой степени индивидуализирована и выражает потребности, особенности, вкусы и привычки данной особи. Животные, как правило, более активны и истребляют обычно большое количество других организмов. Однако даже наиболее пассивные виды живых существ оказывают значительное влияние на внешнюю среду и при отсутствии сопротивления своей деятельности быстро могли бы изменить весь состав биогеоценоза (что иногда и наблюдается). Всей своей жизнедеятельностью особи любого вида организмов воздействуют на биогеоценоз и тем самым «информируют» его о состоянии своей популяции. Свойства особи (фенотипа) данной популяции проявляются в формах ее активности (жизнедеятельности). Однако они бывают отмечены и внешними признаками, характеризующими особей данного вида и также в значительной мере индивидуализированными. Эти внешние признаки играют также известную роль как средства информации, передаваемой от особи к особи той же популяции, а иногда и к особям других видов. Эта информация входит в понятие обратной информации и подлежит такому же контролю и преобразованию в биогеоценозе.
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 573; Нарушение авторских прав?; Мы поможем в написании вашей работы!