КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Возникновение приспособлений (адаптациогенез) 1 страница
|
|
|
|
А. ФИЗИОЛОГИЧЕСКИЕ АДАПТАЦИИ
Обилие и большое значение для жизненной стойкости организма малых физиологических мутаций способствуют, очевидно, тому, что географическая, а частью и экологическая, дифференциация, начинается с небольших различий физиологического характера. Мы уже отмечали, что такие различия в высшей степени характерны не только для невидимых физиологических и для малых мутаций, но и для вполне выраженных морфологических мутаций, являясь обязательным их коррелятом. Это понятно, если мутации по своей природе являются биохимическими изменениями, которые прежде всего ведут к изменениям процессов внутриклеточного обмена веществ и только через это к морфогенетическим преобразованиям. Именно эти элементарные первичные изменения иногда отличаn#1086;рмальный генотип живет при 20о и гибнет при 27° (Banta and Wood, 1927). И отдельные мутации оказываются поэтому в некоторых своих физиологических выражениях благоприятными в обычных условиях культуры. Мутация eversae (D. junebris является при температуре 24–25о более «жизнеспособной» (по проценту вылупления), чем дикая норма (Тимофеев-Ресовский); мутация пурпуровых глаз purple (D. melanogaster) обладает более высокой плодовитостью, а мутация speck связана с большей длительностью жизни. Однако мы знаем, что в некоторых условиях борьбы за существование более низкая плодовитость и даже более короткая жизнь могут приобрести положительное значение.
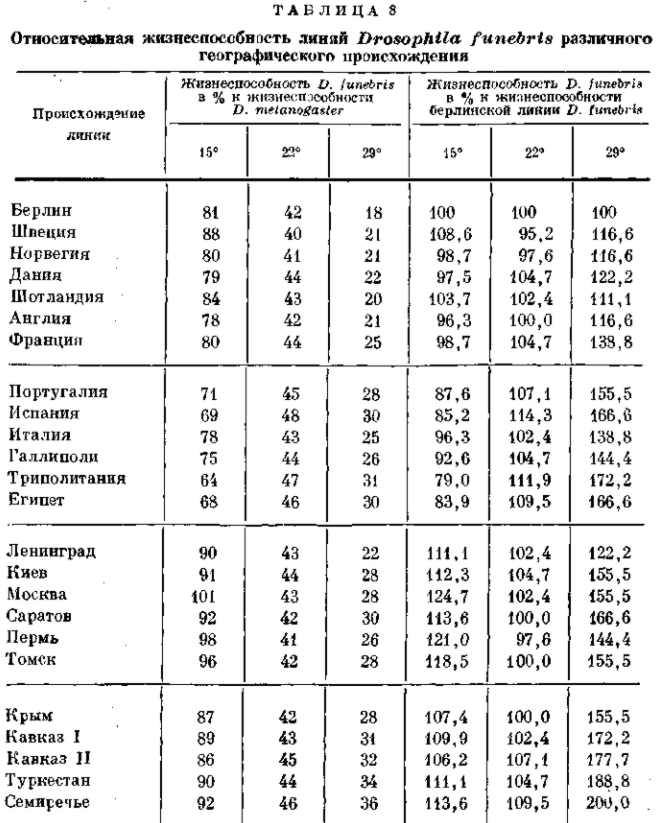
Различия между близкими географическими и экологическими расами и даже между локальными популяциями имеют именно такой характер. Исследования Тимофеева-Ресовского над жизнеспособностью дрозофилы (D. junebris) из различнейших популяций Палеарктики при различных температурах культивирования показали, что юго-западные популяции (из Испании, Италии, Галлиполи, Триполитании и Египта) отличаются большей жизнеспособностью при 29° и меньшей жизнеспособностью при 15° по сравнению с северо-западными популяциями (из окрестностей Берлина, Швеции, Норвегии, Дании, Шотландии, Англии и Франции); северо-восточные популяции (из Ленинграда, Киева, Москвы, Саратова, Перми и Томска) оказались более жизнеспособными как при 15°, так и при 29°, а юго-восточные популяции (из Крыма, Кавказа и Средней Азии) оказались также более жизнеспособными при 15°, но в особенности отличались высокой жизнеспособностью при высоких температурах (29°) (табл. 8). Эти различия находятся в полном согласии с умеренным морским климатом северо-запада, с теплым и мягким климатом юго-запада, с прохладным континентальным климатом северо-востока и с резко континентальным климатом юго-востока. Континентальные популяции отличаются именно высокой стойкостью по отношению к крайним температурам — как низким, так и высоким.
|
|
|
То же самое было найдено Добжанским в отношении североамериканских популяций дрозофилы (D. pseudoobscura). В этом случае изучалась их плодовитость при разных температурах. Оптимальная для плодовитости температура оказалась для более северной и западной расы В ниже, чем для юго-западной расы А К Обычно D. pseudoobscura не выносит температур выше 27,5°. Однако в штате Аризона в окрестностях Юмы были найдены две линии, относящиеся к расе Л, которые удалось культивировать в лаборатории при 28,5–39°. Местность, в которой они были найдены, отличается исключительно жарким летом. Морфологически эти расы неразличимы. У жужелиц Carabus nemoralis географические формы ясно различаются по предпочитаемой температуре.
Чувствительность к разным температурам лежит нередко в основе расовых различий у рыб. «Озимые» и «яровые» расы проходных и полуироходных рыб начинают свои миграции именно при разных температурах воды (Л. Берг, 1934); яровые поднимаются в реки при более высокой температуре и не идут так далеко вверх по течению. По Ильину (1926), географические расы горностаевых кроликов обладают весьма различным температурным порогом образования черного пигмента (сравнивались московская, польская и две германские расы, из них московская обладает наиболее низким, а германские наиболее высоким верхним порогом пигментооб-разования). У многих млекопитающих обнаруживаются расовые различия в предпочитаемых температурах. В экспериментальных условиях, с наличием температурного градиента, серая мышь предпочитала 37,3°, а альбиносы — 34,6° (Гортер). Соответственно и температура их тела различна (36,96° и 33,87°).
|
|
|
Установлены и многие другие расовые различия (а также различия между разными линиями): стойкость к ядам, к паразитарным заболеваниям, иммунитет, морозоустойчивость и засухоустойчивость растений и многие др. Как правило, все эти различия оказываются приспособительными в данных жизненных условиях. В расовых различиях индивидуального развития растений большую роль играют условия влажности и температуры (яровые и озимые расы с дальнейшими различиями) и длина светового дня. Те же факторы играют большую роль в половом цикле птиц и могут быть основой расовых различий. Расхождение во времени цветения у растений и времени полового размножения у животных покоится иногда на очень небольших различиях в физиологических реакциях на изменения во внешних факторах, и, однако, они оказываются часто приспособительными и имеют огромное значение в расо- и видообразовании, так как ведут к половой изоляции новых от исходных форм. Мы в этой связи упоминали о различиях во времени размножения близких рас бабочек (например, дубового шелкопряда), различных рас рыб (сельдей и проходных рыб) и амфибий. То же самое касается, однако, птиц, млекопитающих и очень многих других животных. У млекопитающих имеются нередко большие расовые различия в их поведении, в общей активности и в суточном поведении (дневная, ночная и т. д.). Такие физиологические различия имеют также приспособительное значение; они связаны с местными условиями добывания пищи и защиты от хищников (см. работы Калабухова, Наумова и др.). Все это — определенные экологические различия, которые, как видно, нередко имеют в первую очередь физиологический характер. Это касается и расхождения в питании животных, в формах заботы о потомстве и мн. др.
|
|
|
Равнинные и горные формы живородящей ящерицы (по Рихтеру, 1933), а также лесной мыши (по Калабухову, 1937) обладают наследственными различиями в количестве эритроцитов и в содержании гемоглобина в крови. По Н. Калабухову (1939), северные формы лесной (Apodemus sylvaticus) и желтогорлой (A. flavicollis) мыши отличаются от южных более интенсивным обменом (потреблением кислорода), большей активностью и более низкой предпочитаемой температурой. Близкие и морфологически не отличимые, локальные формы улиток Lymnaea columella из двух разных водоемов (в Пенсильвании, по Бейли, 1939) отличаются по длительности жизни, плодовитости и скорости роста. Гусеницы двух морфологически не различимых рас бабочки Hyponomeuta padella отличаются тем, что одна из них охотнее откладывает яйца на яблоне (90,2%), а другая —на листьях шиповника (79,3%). Очень много фактов такого рода собрано в сводке Калабухова (1941а). Для экологических форм растений физиологические различия не менее характерны, чем для животных. В особенности это касается таких признаков, как засухоустойчивость, устойчивость по отношению к инфекциям и грибным заболеваниям. Экотипы Турессона различаются также по своей физиологической характеристике. Последняя тесно связывается и с морфологическими различиями (см. сводки Розановой, 1940, и Синской, 1948).
Б. ПРИСПОСОБИТЕЛЬНАЯ ОКРАСКА
Пигментация животных по своему развитию и изменчивости приближается к физиологическим признакам. Различия в пигментации также очень тесно коррелированы с физиологическими различиями. В особенности это касается альбиносов, которые обычно отличаются более вялым обменом (хотя белые крысы развиваются, растут и достигают половой зрелости раньше серых) и меньшей активностью. Это верно, однако, и по отношению к меланистиче-ским формам, которые также, по-видимому, обычно обладают и физиологическими различиями. Нас интересует здесь, однако, не этот вопрос, а приспособительное значение расовых различий в окраске. И здесь относительная простота таких изменений ведет к тому, что самые различные уклонения могут оказаться благоприятными в тех или иных частных условиях. Большая или меньшая интенсивность окраски может иметь защитное значение в соответствующих условиях общего фона и освещения.
|
|
|
Распространение меланистической формы хомяка приурочено к влажньщ районам лесостепи на Украине и в Башкирии (С. Гершензон, 1946). Быть может, здесь темная окраска дает ему большую защиту, чем в открытой степи. Однако С. Гершензону.не удалось установить фактов различной истребляемости обеих форм хищниками. Расселение меланистической формы лимитируется зимней гибелью, и это заставляет думать о значении каких-то физиологических коррелятов (в осенних популяциях процент меланистов возрастает либо в результате большего выживания в течение лета, либо в силу большей плодовитости; в весенних популяциях процент меланистов ниже вследствие большей зимней смертности). Здесь защитное значение окраски не доказано. Однако вариации в окраске весьма распространены и во многих случаях, несомненно, приобретают защитное значение (а в других случаях — значение распознавательных отметин, как, например, у многих пауков и у птиц).
Так, например, расовые различия в окраске шерсти североамериканской мыши Peromyscus polionotus, живущей во Флориде, имеют, по Сёмнеру, защитное значение. Обычная серая окраска P. polionotus polionotus под цвет почвы полей заменяется на песчаном побережье очень светлой окраской P. polionotus albifrons. На маленьком полуострове имеется третий подвид P. polionotus leucocephalis, еще более светлый, вполне подходящий к белому цвету песчаной поверхности полуострова. Эти различия оказались наследственными. То же самое было найдено относительно различий в окраске ряда других грызунов, в частности — рода Perogna-thus, которые, по Бенсону, имеют также приспособительный под цвет почвы характер. На песчаной почве распространены светлоокрашенные, иногда почти белые локальные расы этих грызунов, а на темных лавах — темноокрашенные географические или экологические расы.
Наконец, в известных исследованиях Гаррисона (Harrison, 1920) был показан механизм самого возникновения различий в окраске в двух популяциях бабочек ОрогаЫа autumnata, возникших из одной сплошной популяции при разделении леса широкой просекой. В той части леса, где сосна была замещена березой, естественный отбор (преобладающее истребление птицами более темных особей) привел к значительному посветлению популяции бабочек. Различия в окраске географических и экологических рас имеют, как сказано, наследственный характер и покоятся на комбинациях мутаций, которые иногда производят впечатление очень простых различий в отдельных факторах (например, различные формы богомолов или различные мимикрирующие формы самок Papilio polytes и Р, darclanus).
В. ИНДИФФЕРЕНТНЫЕ МОРФОЛОГИЧЕСКИЕ РАЗЛИЧИЯ
Систематики постоянно подчеркивают, что расовые и видовые различия имеют главным образом индифферентный характер и не могут рассматриваться как приспособления. К таким утверждениям следует всегда относиться с осторожностью, так как мы в огромном большинстве случаев не знаем экологических и особенно биоценотических соотношений с такой полнотой, чтобы иметь правильное суждение о физиологическом и биологическом значении тех или иных признаков. Тем не менее, если бы это было и так, в этом нельзя усмотреть затруднений для теории естественного отбора, против которой обычно такие аргументы выдвигаются.
Прежде всего отметим, что положительная роль естественного отбора основывается всегда на обратном процессе — избирательной элиминации менее приспособленных особей. Поэтому вместе с накоплением благоприятных изменений идет всегда и накопление безразличных мутаций, малых мутаций, а также и условно вредных и частично вредных мутаций. Вредные выражения мутаций нейтрализуются в процессе стабилизирующего отбора модификаторов, а безразличные выражения могут остаться в качестве признаков рас и даже видов. Они могут быть использованы и в качестве распознавательных отметин (приобретают тогда положительное значение в процессе полового размножения, в уходе за потомством и в стадной жизни).
Даже такой совершенно безразличный морфологический признак, как правое или левое вращение улиток, оказался связанным с физиологическими различиями (Гаузе, Смарагдова, 1939). Улитки (Fruticicola lantzi) с левозавитыми раковинами в лабораторных условиях быстро падали в весе, по всей вероятности, вследствие более интенсивного обмена. Между тем, в Казахстане в некоторых местах (в окрестностях Алма-Ата) левозавитые раковины составляют громадное большинство популяций (в других местностях полностью господствуют правозавитые улитки).
Близкие виды постоянно отличаются по плодовитости, скорости созревания, темпу размножения, по времени спаривания и размножения, по питанию, характеру активности и ее распределению в дневное и ночное время. Исследование всегда показывает связь расовых, подвидовых и видовых различий с особенностями местообитания и жизни в определенной биоценотической обстановке. Неизменно эти различия оказываются приспособительными.
Современные исследования экологов дают огромный материал для физиологической оценки различий между близкими видами, подвидами и даже отдельными популяциями одного и того же вида или подвида. В этом отношении большое значение имеют прекрасные работы Калабухова по изучению близких форм грызунов, которые ясно показывают адаптивность этих различий. Весьма возможно, что во многих случаях физиологическая адаптация действительно предшествует морфологической (Промптов, 1933). Однако вряд ли это составляет общее правило. Детальные исследования морфологических признаков вскрывают и здесь адаптивность подвидовых и расовых различий. Напомню результаты исследований окраски экологических рас и подвидов различных грызунов, проведенных рядом американских авторов (Sumner, Dice, Benson и др.). Эти исследования показали защитный характер окраски даже отдельных популяций — от почти белой на песчаной почве до почти черной на темных лавах. Сошлюсь также на исследования видов и подвидов рода Dipodomys (Grinell I., 1922), показавшие приспособленность к бегу скачками в определенных условиях открытых степей (более длинные конечности) или кустарных зарослей (конечности короче и толще), а также на исследования видов и подвидов рода Passerella (Linsdale I., 1928) в отношении развития скелетных частей крыла, плечевого пояса и грудины в связи с областью гнездования и длиной пути их перелетов на юг (развитие этих частей тем выше, чем длиннее пути миграции).
Во всех тех случаях, когда экологи производят детальное исследование, вскрывается адаптивный характер не только физиологических, но и морфологических видовых различий. Почему же все-таки систематики так усиленно подчеркивают «индифферентный» характер видовых различий? Это объясняется именно тем, что адаптивные видовые различия неудобны для систематика — они не всегда видны на консервированном материале (физиологические различия) или требуют более сложного анатомического исследования («организационные» различия), и притом обычно — ввиду трансгрессивного их характера — большого материала и биометрической его обработки. Систематики идут по этому последнему пути; однако и этот путь дает возможность установить адаптивный характер различий лишь при наличии исчерпывающих наблюдений над жизнью данного организма в его естественной обстановке. Следует учесть и еще одно обстоятельство, когда сравнивают адаптивность видовых различий с адаптивностью признаков более крупных таксономических подразделений.
Конкретным объектом эволюции являются всегда только виды с их видовыми различиями. Виды превращаются в другие виды, дифференцируются и дают начало более или менее многочисленным новым видам. При всем этом виды всегда остаются видами. Признаки организации меняются в связи с изменением соотношений с элементами внешней среды, и какие из этих изменений будут иметь ограниченный характер «видовых» различий, а какие в дальнейшем процессе эволюции приобретут более общий характер родовых, порядковых или классовых различий,— заранее предсказать невозможно. Только ретроспективно -мы можем сказать, что такие-то различия, возникнув когда-то в качестве видовых, приобрели затем значение признаков, характеризующих семейство или другое крупное подразделение. Одни признаки имеют кратковременное существование — они связаны с ограниченными условиями жизни данного вида и исчезают вместе с его преобразованием в другой вид, живущий при иных условиях. Другие признаки имеют более длительное существование — они сохраняют свое значение и при некотором изменении внешней обстановки и, следовательно, при распаде вида на целую группу видов. Естественно, что более длительное время сохраняют свое значение признаки явно адаптивного характера. Признаки наиболее общие именно потому и входят в характеристику крупных таксономических подразделений, что их адаптивный характер не теряется с изменением локальных — географических или экологических — условий существования (хотя в деталях они, конечно, изменяются). Маловажность других различий, которые могут иметь весьма ограниченное значение в данной местности, в данных экологических условиях или для данной популяции (например, распознавательные отметины), является причиной их легкой изменяемости и сравнительно кратковременного существования. Маловажность признаков затрудняет вскрытие их адаптивности (без специального исследования в естественной обстановке), а с другой стороны, она ограничивает их значение пределами низших систематических подразделений. Систематики, подчеркивающие широкий, приспособительный характер признаков больших подразделений и нейтральный характер видовых различий, попадают, следовательно, в логический круг. Различия ограниченного значения, т. е. «нейтральные» признаки, потому и избираются для видовой диагностики, что они резко изменяются вместе с изменением самого вида. Признаки гораздо более важные — «организационные» — меняются медленнее, и их преобразования, нередко количественные, гораздо менее удобны для систематика. Мы не раз подчеркивали ведущее значение центральной нервной системы в эволюции высших позвоночных. Определяя характерное поведение данного вида, подвида и даже экологической расы животных, видовые различия в строении центральной нервной системы имеют, несомненно, адаптивное значение. Вряд ли, однако, было бы удобно класть в основу видовой диагностики специфику микроскопической архитектоники головного мозга.
Г. МОРФОЛОГИЧЕСКИЕ АДАПТАЦИИ
Морфологические выражения мутаций наиболее изучены. Однако как раз здесь трудно установить существование положительных изменений, которые могли бы иметь значение адаптации. Приходится признать, что морфологические признаки, будучи конечными звеньями в цепи биохимических и физиологических реакций и основанные всегда на сложных онтогенетических взаимозависимостях, чаще всего испытывают очень грубые изменения. Поэтому морфологические адаптации могут строиться только на очень малых мутациях, не имеющих значительного деструктивного характера.
Нетрудно, конечно, понять явления редукции органов, потерявших свое значение, так как большинство мутаций связано именно с явлениями недоразвития. Такие адаптивные преобразования, как рудиментация крыльев у островных насекомых, редукция глаз у пещерных и роющих животных или редукция конечностей у змееобразных позвоночных, настолько ясны, что на этом можно не останавливаться (Шмальгаузен, 1938а, 1942).
Не представляет особых трудностей и вопрос о количественных изменениях, лежащих в основе прогрессивного развития органов. Форма, число и длина щетинок у дрозофилы, форма и длина конечностей и антенн, число омматидий в глазах, число яйцевых трубок и яйцевых камер — все эти величины меняются во многих весьма обычных мутациях. В основе новообразований могут также лежать и чисто количественные изменения. К числу весьма распространенных мутаций относятся всевозможные удвоения, ведущие к увеличению числа общегомологичных органов — удвоение щетинок у дрозофилы, удвоение конечностей. У позвоночных бывают частичные удвоения конечностей (полидактилия у кур, голубей; также у млекопитающих), увеличение числа сегментов тела и т. п. Такие изменения лежат, очевидно, в основе явлений «полимеризации», которые могут вести к новым, более разнообразным дифференцировкам между общегомологичными органами. В процессе эволюции такие изменения играют, очевидно, немалую роль.
Гораздо труднее проследить за начальным возникновением качественно новых адаптации. Фактических наблюдений этого рода имеется пока еще очень мало. Наиболее вероятным представляется мне предположение, что такие совершенно новые адаптации возникают именно на базе тех «нейтральных» различий, которые в большом числе накапливаются в составе «резерва» внутривидовой изменчивости и в процессах перекомбинирования мутаций непрерывно меняют свое выражение. При изменениях внешней среды или при миграции самого организма могут сложиться и новые, более благоприятные соотношения между организмом и средой.
Изменения в жилковании крыла принадлежат к наиболее распространенным выражениям разнообразных мутаций дрозофилы. Вместе с тем тип жилкования представляет важный и довольно устойчивый систематический признак, характеризующий целые группы насекомых и, очевидно, не лишенный приспособительного значения (он определяет упругие свойства крыла как летательного органа). Некоторые крыловые мутации все же распространяются в популяции и могут послужить материалом для приспособительных преобразований конструкции крыльев. У видов рода Andrena (Hymenoptera) встречается мутация second cubital crossvein. У A. albicans она наблюдается очень редко, у А. ргаесох и особенно у A. vaga — несколько чаще. У A. serica эта мутация довольно обычна (около 8% особей), а у A. argentata встречается еще чаще (около 20% особей). Очевидно, эта мутация приобретает уже какое-то положительное значение. Наконец, у A. neglecta это изменение стало нормальным признаком вида (Zimmermann К., 1933). Качественный характер имеет и мутация simplex полевки Microtus arvalis, выражающаяся в изменении строения коренных зубов. Эта мутация распространяется в Шлезвиг-Гольштинии, достигая там значительной концентрации — выше 85% особей, что указывает на ее положительное значение в данных условиях.
Если мы говорили раньше о частично вредных или обезвреженных морфологических коррелятах физиологических мутаций и возникающих на их базе физиологических адаптации, то теперь мы должны подчеркнуть, что и морфологические изменения обладают не только физиологическим выражением, но и своими коррелятивными связями. Если добавочные физиологические и морфологические (плейотропные) выражения морфологических мутаций не слишком неблагоприятны (снижение «жизнеспособности» и «плодовитости»), то они нейтрализуются в процессе отбора наиболее благоприятных комбинаций. Тем самым они теряют свои второстепенные выражения. Однако всякий морфологический признак играет какую-то роль в жизни организма. Поэтому, входя в состав приспособленной нормы, морфологическое изменение всегда приобретает если не физиологическое, то биологическое или экологическое значение.
Физиологическое выражение морфологических адаптации состоит в характерных для них жизненных функциях организма. Так, даже изменение величины тела может иметь физиологическое значение. Увеличение размеров тела связано всегда с более экономным обменом веществ, так как продукция энергии определяется объемом живого вещества или активных тканей организма, а трата энергии — поверхностью организма. Это касается всех животных, а не только гомойотермных. Пойкило-термные животные лишь непрерывно растрачивают продуцируемую тепловую энергию, отдавая ее в окружающую среду, а го-мойотермные животные частично сберегают, используют и поддерживают температуру на оптимальном уровне. Поэтому «правило Бергмана» о возрастании величины тела (близких групп животных) при переходе от теплого климата к более холодному частично распространяется и на «холоднокровных» животных. Однако значительная интенсивность обмена (и жизнедеятельности) у птиц и у млекопитающих естественно ставит вопрос о потере энергии через теплоотдачу более остро, чем у других животных. Поэтому правило Бергмана имеет здесь более всеобщий характер. Большую величину тела у птиц и млекопитающих холодных стран мы должны считать морфологическим признаком большого физиологического значения. То же самое относится к таким характеристикам, как величина ушей у некоторых млекопитающих и длина хвоста у мышей. Во многих случаях они имеют прямое отношение к терморегуляции (как части тела с менее развитым волосяным покровом и богато развитой сетью кожных кровеносных сосудов). Большие уши южных лисиц способствуют теплоотдаче и предохраняют тело от перегревания, а малые уши песца, наоборот, сокращают возможную потерю тепла. Длинные хвосты южных мышей и короткие хвосты северных имеют такое же физиологическое значение.
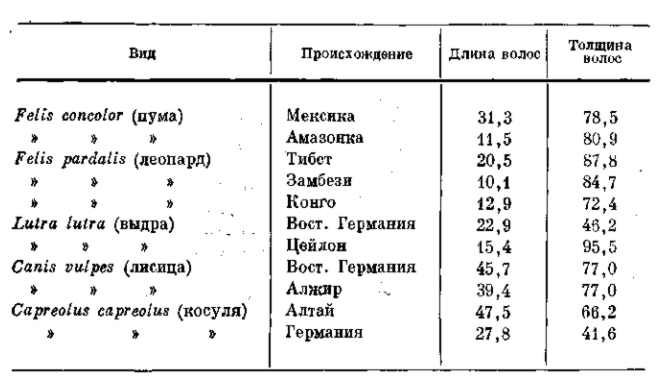
Чисто морфологические различия в числе, длине и толщине волос у млекопитающих играют такую же роль в регулировании теплоотдачи. Эти изменения исключительно просты, и вместе с тем их адаптивность совершенно ясна. Приводим таблицу 9, показывающую различия в длине и толщине волос у северных и южных млекопитающих.
Интересную попытку проследить возникновение морфологических адаптации определенного функционального значения сделал Н. В. Дубовский (1941) в своем сравнительном и экспериментальном исследовании органов движения и прикрепления у ракушковых рачков (Ostracoda). Эти рачки плавают при помощи двух пар антенн, снабженных длинными гребными щетинками, и ползают, опираясь на вторую пару ножек, снабженных конечными коготками (у морских форм для хождения используются три пары ножек).
Формы, исключительно ползающие, характеризуются укорочением гребных щетинок первой пары антенн, рудиментацией или даже отсутствием щетинок второй пары антенн; укороченными, утолщенными и более мускулистыми конечностями, особенно у форм, зарывающихся в песчаный грунт. В пределах многих родов пресноводных плавающих остракод имеются отдельные виды, приспособленные исключительно для ползания. Эти последние характерны для текучих вод и, очевидно, произошли от исходных форм стоячих водоемов.
Для экспериментального исследования избран типичный представитель речных Ostracoda (Волги и Сев. Донца) — Cypria curvifurcata, У этого вида установлено существование различий между локальными формами, заселяющими места реки с песчаным дном и более сильным течением, и формами, заселяющими участки с заиленным дном и медленным течением. Биометрическая обработка материала, полученного в результате многочисленных измерений, показала существование вполне достоверных различий по очень многим показателям. В общем формы, живущие на песчаном дне участков рек с более сильным течением, имеют конечности и их придатки (коготки и щетинки) сравнительно короткие, толстые, сильнее хитинизированные, очевидно, приспособленные к передвижению на крупнозернистом песке, к закапыванию в нем и к укреплению и противостоянию более сильному течению. Формы с заиленных мест рек со слабым течением характеризуются сильным развитием придатков конечностей (длинные и тонкие коготки и щетинки) и большей длиной самих конечностей. Длинные конечности характерны и для лазающих по растениям представителей Cypridae из стоячих пресных водоемов. Сравнительно-морфологический анализ морских Ostracoda (сем. Cytheridae) также показывает, что длинные конечности с тонкими придатками имеются у видов, живущих на более зыбком илистом грунте. Эксперименты поставлены следующим образом. Особи, взятые с заиленных мест реки, помещались в деревянный лоток вместе с некоторым количеством грунта и подвергались действию тока воды, который смывал животных в подставленный сачок. Оставшиеся животные смывались затем более сильной струей воды в другой сачок. Смытые вначале животные первого сачка сравнивались с оставшимися животными из второго сачка (после фиксирования, измерения и биометрической обработки полученных величин). По 12 признакам получены статистически достоверные различия между смытыми и несмытыми особями. Именно у несмытых особей оказались более короткие коготки второй антенны, более короткие дистальный членик и коготок первой ножки и более короткие основание и коготки фурки. По этим признакам несмытые особи приближаются к локальным формам, характерным для мест с более быстрым течением. Это показывает приспособительный характер изменений и эффективность отбора по перечисленным признакам в экспериментальных условиях, причем направление этого отбора совпадает с направлением изменений в естественных биотопах.
ТАБЛИЦА 9. Длина (в мм) и толщина (в мк) волос географических рас, обитающих в разных климатических поясах (по Rensch, 1929)
При изучении морских Ostracoda из сем. Cytheridae найдены подобные же различия между прибрежными формами, живущими в зоне прибоя, цепляющимися за водоросли и обладающими наиболее короткими члениками конечностей и коготками, формами, ползающими на песчаном грунте, и, наконец, формами, живущими на илистом дне. Эти последние обладают наиболее длинными конечностями, щетинками и коготками, приспособленными не столько для цеплянья и лазанья, сколько для того, чтобы при помощи увеличенной поверхности держаться на зыбком иле, а также зарываться в него.
Экологическое значение морфологических изменений определяется именно положением организма во внешней среде и, в особенности, его соотношениями с другими организмами. Таковы уже упомянутые явления защитной окраски, таковы же и различные другие средства пассивной (а также и активной) защиты. Хитиновые, роговые и костные наружные панцири с шипами и иглами могли развиваться на базе самых незначительных и очень простых изменений адаптивного характера (в смысле относительной эффективной защиты от хищников или паразитов).
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 545; Нарушение авторских прав?; Мы поможем в написании вашей работы!