КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Доминантность. Баланс нормального генотипа обычно достаточно защищен от нарушений со стороны небольших мутаций. В гетерозиготе это выражаетсяв доминировании нормы. 1 страница
|
|
|
|
Так как многочисленные мутации распространяются в популяциях лишь в гетерозиготном состоянии, то практически в борьбе за существование конкурируют всегда самые разнообразные гетерозиготы. В естественном отборе преимущества в большинстве случаев окажутся на стороне уже приспособленной «нормы». Погашение выражения различных неблагоприятных мутаций в гетеро-зиготе, т. е. усиление доминирования «нормы», достигается, как мы видели, отбором наиболее благоприятных генных сочетаний. В результате создается известная защита нормального формообразования от возможных нарушений, создается известный «фактор безопасности» (Haldane, 1929). Генетики отмечают, что в этом случае активность нормального «гена» достигает оптимального уровня, так что его удвоение в гомозиготе уже не вносит дальнейших изменений. Мы уже рассматривали факты обезвреживания мутаций в гетерозиготе (через развитие их рецессивности), а тем самым разобрали и вопрос об эволюции доминантности нормы. Мы видели, что различные авторы представляют себе эти процессы по-разному, и пришли к выводу, что в основе этих разногласий лежат вполне объективные различия конкретных путей, при помощи которых в эволюции достигаются сходные результаты. Доминирование нормы представляет во всяком случае побочный результат развития более устойчивых форм. Однако на основе этой устойчивости нормального формообразования лежат не только генетические механизмы, но и более сложные механизмы индивидуального развития, имеющие также до известной степени регуляторный характер.
Б. ФЕНОГЕНЕТИЧЕСКИЕ1 СИСТЕМЫ
1 В личном экземпляре И. И. Шмальгаузена — «эпигенетические» (Waddington). — Ред.
Основным источником преобразования системы взаимозависимостей частей развивающегося организма служат, очевидно, элементарные изменения взаимосвязи, обнаруживаемые в отдельных мутациях. Они выражаются в известных явлениях плейотропизма. Общая характеристика явлений была дана ранее (см. 1–4, Б).
|
|
|
Первично, очевидно, любое наследственное изменение должно было изменять весь организм в целом. Если те или иные части этого организма находятся в разном положении относительно факторов среды, т. е. процессы обмена в них идут несколько различно, то, очевидно, изменение одного наследственного фактора приведет к различным изменениям в этих, разных по своей физиологической ситуации, частях. Если эти изменения имеют, при данных условиях, необратимый для особи характер, т. е. касаются образования более стойких продуктов, то возможна на этом пути наследственная дифференциация организма, определяемая, однако, в своей реализации, на известной стадии развития, некоторыми факторами внешней среды. Дифференциация возможна и в том случае, если детерминирующие ее продукты остаются внутри тех клеток, специфический характер обмена которых определяет специфику действия данных продуктов.
По мере усложнения строения организма происходит известная дифференциация детерминирующих продуктов. Каждый из этих продуктов действует различно на различные части, но их действие всегда ограничивается теми клетками, в которых эти продукты образуются.
Эту форму плейотропного действия можно обозначить как множественное, или полиморфное, выражение мутации. Изменение одного гена сказывается так или иначе на всех клетках организма. В зависимости от уже имеющейся в наличии дифференцировки, от положения и, следовательно, характера обмена в разных клетках изменение преобретает разный характер и ведет к различным изменениям продуктов дифференцировки. Взаимозависимости в изменениях различных частей, определяемые здесь изменениями в первичных же морфогенных продуктах, мы обозначаем как геномные и, в частности, феногенетические корреляции.
|
|
|
Однако совершенно невероятно, чтобы изменение одного гена, ведущее к благоприятному изменению в одной части, привело к благоприятному изменению в другой части (А. А. Малиновский). Следовательно, эта форма плейотропизма, характерная для мозаичного организма, но широко распространенная и при регуляционном типе развития, сама по себе не может приобрести положительного значения в процессе эволюции. Путем непрерывного отбора других, особенно малых, мутаций происходит такое их комбинирование, которое приводит к погашению всех неблагоприятных прояв-. лений плейотропизма, т. е. к уничтожению большинства видимых выражений данной мутации. Так как, кроме одного основного проявления (представляющего собой предмет отбора), все другие, как правило, оказываются неблагоприятными, то эволюция идет все время в сторону устранения плейотропного выражения мутаций и выработки их специфически локального действия, на основе взаимодействия между различными внутриклеточными процессами, зависящими от изменения многих генов. Лишь в редких случаях, в результате отбора благоприятных комбинаций, возможно установление нескольких положительных выражений одной мутации (в сочетании с рядом модификаторов). Как правило, положительное значение в процессе эволюции могут приобрести лишь взаимозависимости, определяемые целым комплексом таких изменений. Такие «морфогенетические» корреляции выражаются уже не в явлениях обычного плейотропизма и методами генетического анализа не могут быть вскрыты.
Наиболее резко выражено локальное действие морфогенных продуктов у мозаичных организмов и, в частности, у дрозофилы. Что это действие в самом деле ограничивается теми клетками, в которых эти продукты образуются, видно по мозаикам и отдельным соматическим мутациям, в которых изменение сказывается в резко ограниченной области, простирающейся иногда (IB случае позднего возникновения) на очень небольшой участок — при изменении окраски глаза, например, на несколько фасеток.
Возможна, однако, и другая форма плейотропизма, которую можно обозначить как многостепенное или полифазное выражение мутации. В этом случае морфогенные продукты распространяются путем нередко весьма ограниченной, но иногда и более свободной диффузии, а специфика морфогенетической реакции зависит не только от этих продуктов, но и от условий их распространения в теле, а также, конечно, в значительной мере и от уровня диф-ференцировки, положения и, следовательно, характера обмена в клетках различных частей развивающегося организма. Число возможных дифференцировок, достигаемых в результате действия одних и тех же морфогенных продуктов, при этом значительно возрастает вследствие увеличения числа комбинаций действия различных веществ, достигающих разной концентрации в тканях различной ситуации, разного возраста и дифференцировки (а следовательно и обмена). Такие взаимозависимости можно рассматривать как элементарные формы морфогенетических корреляций.
|
|
|
Диффундирование морфогенных продуктов (в виде «геногормонов») доказано в некоторых случаях и для мозаичных организмов. Однако гораздо большее значение оно приобретает при регуляционном типе развития, как это в особенности характерно для позвоночных животных.
Уже Стертевантом (1929а) было доказано существование у дрозофилы выделяемого яичником гормона, который подавляет в нор-, мальной мухе проявление окраски vermilion. To же самое было затем подтверждено опытами Бидла и Эфрусси (1935–1937) с трансплантацией имагинальных дисков глаз, взятых от личинок различного наследственного строения, на личинки иного генотипа. Почти все глазные окраски развивались автономно, т. е. под контролем факторов, заключающихся в клетках самого зачатка. Однако окраски vermilion и cinnabar развивались в зависимости от хозяина и, следовательно, в теле нормального хозяина (и большинства других мутантов) не проявлялись. Опыты трансплантации показали также возможность обратного влияния трансплантата на ткани мутантного хозяина. Наконец, опытами инъекции лимфы показано, что действительно вещества порядка «геногормонов» распространяются в этом случае путем диффузии по всему телу (Эфрусси, Кланси и Бидл, 1936). То же самое показывают и опыты кормления личинок дрозофилы одного генотипа кашицей из личинок или куколок другого генотипа, содержащих такие диффундирующие вещества, действующие на окраску глаз (Бидл, Эфрусси, 1936, 1938; Нейгауз, 1939, 1940). Кроме яичника, местом образования этих «геногормонов» оказались также жировое тело и мальпигиевы сосуды.
|
|
|
Очень ясную картину гормонального влияния дали исследования Кюна и его сотрудников над развитием амбарной огнёвки Ephestia kuhniella. Здесь показана особо сложная цепь зависимостей пигментообразования в разных частях тела и в разных возрастах. Изменения вызываются и здесь «геногормонами», вырабатываемыми главным образом в половых железах, именно в семеннике (а отчасти и в центральной нервной системе), откуда они распространяются по всему телу. Изменение сводится в основном к изменению скорости пигментообразования и сказывается на окраске покровов гусеницы, ее ложных глазков, оптического ганглия и на глазах взрослой моли.
У позвоночных формообразовательное значение гормонов общеизвестно. Изучено также значение продуктов, действующих на небольшие расстояния и известных под названием индукторов (в широком смысле слова). Связь с отдельными «генами» здесь в большинстве случаев не установлена, так как не производилось сравнительного исследования действия индукторов у форм, отличающихся одним или немногими «генами». Исключение составляют лишь немногие работы, как, например, феногенети-ческий анализ куроперости, проведенный Пеннетом и Бейли (1921), феногенетический анализ карликовости у мышей по исследованиям Смита и Мак-Доуелла (1931). В первом случае изменение зависит от изменения реактивности тканей на женский половой гормон, во втором случае — от изменения продукции ростового гормона в гипофизе. К этому можно еще добавить исследования Бонневи (1934) над аномальными мышами Литтля и Багга. Здесь причиной развивающихся дефектов оказалось действие цереброспинальной жидкости, в избытке выступавшей из IV желудочка мозга и собиравшейся в различных местах под кожей, в особенности у глаз или в конечностях.
Я проанализировал у кур развитие некоторых расовых признаков, имеющих характер простых мутаций, т. е. зависящих в своей реализации от изменений в отдельных генах. Приведу несколько примеров расовых признаков со связями возрастающей сложности.
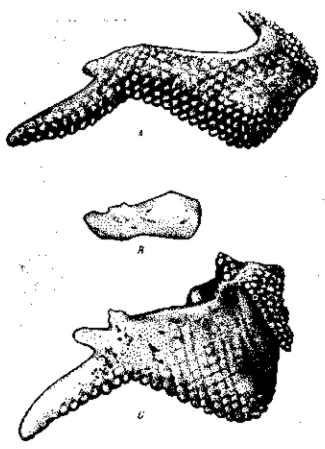
Рис. 34. Закладка лишнего пальца в крыле зародыша полидактильной курицы (фавероль) и ее регуляция. А —крыло 9-дневного зародыша обыкновенной курицы; В — крыло с раздвоенным зачатком первого пальца 7-дневного зародыша фавероли; С — крыло 9-дневного зародыша фавероль. На более поздних стадиях зачаток останавливается в своем развитии, получает очень изменчивое выражение и совершенно редуцируется.
Полидактилия кур выражается в частичном удвоении зачатка конечности по преаксиальному ее краю, в зеркальном расположении, как это наблюдается и в экспериментальных ДЕойниках. Закладываются 2–3 лишних пальца, но вполне развивается лишь один добавочный палец, дифференцирующийся по типу второго пальца. Закладка происходит и в задней и в передней конечностях, но в передней ее развитие затем подавляется (рис. 34). Следовательно, плейотропный эффект этой мутации вторично уничтожается. Здесь имеется плейотропизм первого рода, т. е. типичное множественное выражение изменения гена.
Мохноногость кур выражается у эмбриона очень ранней концентрацией мезенхимы под кожей постаксиального края задней конечности, сходной с нормальной ее концентрацией в передней конечности (соответственно закладке маховых перьев). Можно думать о восстановлении ллейотропного эффекта, который в истории птиц был полностью подавлен как вредный (здесь, естественно, вспоминается гипотетический Tetrapteryx, бывший, по Бибу, переходной формой от лазающих псевдозухнй к перво-птяцам). Концентрация мезенхимы под кожей ведет к нарушению нормального расположения зачатков чешуи на цевке, а затем к закладке ряда перьев, напоминающих маховые (рис. 35). Это, видимо, вторичный эффект изменения гена. Мезенхима, идущая на построение этих закладок, имеет явно скелетогенное происхождение, и в связи с этим более или менее недоразвиваются фаланги 4-го пальца (брахидактилия), палец изгибается на тыльную сторону, и на нем недоразвивается коготь. Об этих изменениях можно говорить как о третичных выражениях мутации мохноногости (именно доминантной мохноногости типа брама и бентамок). Ясно проявляется плейотропизм второго рода, т. е. многостепенное выражение изменения гена.

Рис. 35. Развитие мохноногости у кур. А и В — закладка первого и второго ряда перьев ноги («маховых» и «кроющих»). А — конечность 11-дневного зародыша брама с почти нормальным четвертым пальцем; Б — конечность 11-дневного зародыша брама с резко выраженной брахидактилией и дорсальным изгибом четвертого пальца; С — конечность 11-дневного зародыша бентамки с сильно развитым оперением и резко выраженной брахидактилией (вид с тыльной стороны); Д — конечность 11-дневного зародыша фавероли с зачатками перьев и вполне нормальным четвертым пальцем
Еще более сложная цепь зависимостей обнаруживается при развитии хохла у гуданов. Здесь первое видимое выражение мутации выявляется на эмбриональной стадии у шестидневных зародышей в усиленной секреции цереброспинальной жидкости клетками эпендимы переднего мозга. Образуется масса крупных вакуолей, лежащих непосредственно под эпендимой. В результате сильно раздуваются передние желудочки головного мозга (рис. 36, 37). Мозговая жидкость, очевидно, просачивается через тонкую крышу III желудочка и действует разрушающим образом на перепончатую закладку крыши черепа, полностью ее дезорганизуя (сходное действие цереброспинальной жидкости установлено и в упомянутых исследованиях Бонневи; кроме того, продукция фибринолитических ферментов клетками эпендимы показана Вейссом и в культурах in vitro). Эта дезорганизация зачатка крыши черепа является вторичным выражением хохлатости гуданов. В связи с изменением формы головного мозга стоит своеобразное перемещение зачатков костей черепа к его основанию как ряд третичных изменений (рис. 41, 42). Скелетогенная мезенхима крыши черепа концентрируется под кожей, здесь же разрастается обильная сеть мозговых кровеносных сосудов, и на этом месте развивается масса увеличенных зачатков перьев, образующих затем характерный хохол на голове. Все это можно обозначить как дальнейший ряд третичных и четвертичных изменений. Наконец уже постэмбрионально под кожей развиваются отдельные островки костной ткани, соединяющиеся затем перекладинами и образующие решетчатую вначале, своеобразную структуру— новую, сильно вздутую крышу черепной коробки (рис. 38).
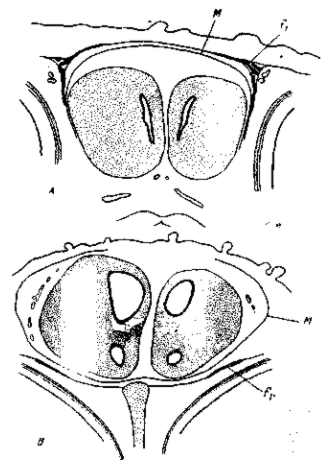
Рис. 36. Поперечные разрезы через переднелобную область головы 10-дневных зародышей нормальной курицы (фавероль) и гидроцефальной (гудан). У гидроцефального зародыша желудочки переднего мозга раздуты, крыша перепончатого черепа разрушена, закладки лобных костей сдвинуты к основанию мозга fr — лобные кости, М — зачаток твердой мозговой оболочки
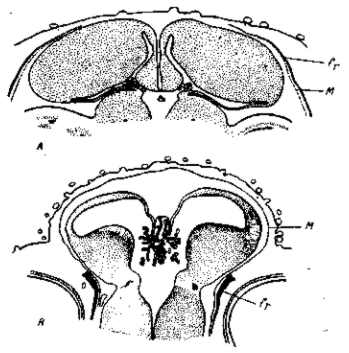
Рис. 37. Поперечные разрезы через лобную часть головы 10-дневных зародышей в области монроевых отверстий мозга. Вверху — фавероль, внизу — гудан. У гидроцефального зародыша желудочки раздуты, лобные кости сдвинуты к основанию переднего мозга (их дорсальные края как будто срезаны). На вздутой верхней поверхности головы видны сильно развитые зачатки перьев (хохол)

Рис. 38. Череп гидроцефальной курицы с решетчатой шишкой в лобной области и для сравнения (внизу) — череп обыкновенной курицы
Поскольку вся эта цепь изменений нормального развития зависит от изменений одного гена, мы здесь можем определенно говорить о плейотропном выражении мутации. Однако этот плейотропизм имеет явно иной характер, чем то, что мы находим у дрозофилы. Если мы там говорили о множественном эффекте, понимая под этим только различную локализацию отдельных изменений, то здесь мы можем говорить о многостепенном, или полифазном, выражении плейотропизма. Несомненно, что и здесь отдельные неблагоприятные выражения могут быть вторично подавлены действием модификаторов. Это доказывается фактом существования хохлатых кур без черепной шишки, а также исследованиями Фишера, установившего, что у японских шелковых кур выражение мутации гидроцефалии подавляется комплексом других факторов, между тем как развитие хохла остается.
Учитывая, что при многостепенном выражении изменения одного гена мы имеем явные зависимости между морфогенетическими процессами, мы можем уже с полным правом говорить о морфогенетических корреляциях.
Многостепенное выражение одной плейотропной мутации можно себе представить как результат последовательной реализации известных реакций. Если первичные морфогенные продукты образовались в самих клетках и их действие сказалось, например, в усилении секреции клеток эпендимы, то эти вторичные продукты секреции диффундировали в ткани и разрушали зачаток крыши черепа, оказав какое-то действие на составлявшие его мезен-химные клетки. Изменение в обмене мезенхимных клеток было связано с изменением их реактивной способности — они переместились под кожу. Выделяемые ими третичные продукты вызвали здесь разрастание кровеносных сосудов, более мощное развитие зачатков перьев и т. д. Это действие можно выразить диаграммой (рис. 39).
Если мы, однако, уже раньше отмечали полную невероятность того, что различные выражения изменения одного гена могут иметь положительное значение для жизни организма в какой-либо конкретной обстановке, то и для этих форм плейотропиз-ма все сказанное сохраняет свою силу. Если одно выражение изменения гена является благоприятным, то другие его выражения скорее всего окажутся неблагоприятными. Эти последние могут быть подавлены в процессе естественного отбора модификаторов. Это видно из следующего. Вторичные результаты изменения гена обычно весьма изменчивы, что само уже указывает на существование модификаторов. Так, например, мутация мохионогости выражается (в зависимости от времени мобилизации скелетной мезенхимы) весьма различной степенью брахидактилии, и в некоторых случаях это последнее выражение может быть полностью подавлено (как, например, у фаверолей). Если в процессе эволюции мохноногость может приобрести положительное значение, то побочное, отрицательное выражение той же мутации — брахидактилия, очевидно, будет подавлено.
Мутация полидактилии выражается в закладке лишнего пальца в обеих парах конечностей, но в крыле ее проявление затем подавляется. При этом изменчивость зачатка лишнего пальца крыла очень велика, что указывает на роль модификаторов в его редукции.
Мутация хохлатости кур могла бы приобрести в процессе эволюции положительное значение (хотя бы как распознавательный признак), но недоразвитие крыши черепа есть во всяком случае очень вредное изменение. И это выражение может быть подавлено, как показывает пример японских шелковых кур. Очевидно, это изменение произошло за счет уменьшения секреции мозговой жидкости и уменьшения реактивности мезенхимы, либо, еще вероятнее, за счет более раннего и свободного выделения мозговой жидкости наружу (чем предотвращалось раздувание желудочков и последующее весьма интенсивное истечение жидкости).
Таким образом, многостепенная форма выражения плейотро-пизма в процессе эволюции неизбежно осложняется изменением морфогенных продуктов, в связи с изменением других генов, что частью осуществляется через более сложные формы взаимодействия с другими тканями (т. е. через вторичные и третичные продукты). Это осложнение морфогенетических процессов может быть выражено приведенной второй диаграммой (рис. 39). Здесь вторичные и третичные (В, С, D) выражения основной мутации видоизменены взаимодействием со вторичными продуктами других клеток, которые нейтрализуют вредные выражения и, быть может, дают новые, благоприятные для жизни организма выражения комплексного действия ряда мутаций.

Рис. 39. Схема многостепенного выражения плейотропии (I), изменяемого в процессе эволюции вследствие естественного отбора модификаторов (II) и дальнейшего усложнения форм взаимодействия (III), в результате которого развивается типичная система морфогенетических корреляций
Пока мы имеем дело с цепью реакций, определяемых одной основной мутацией, вторичные выражения которой видоизменены действием модификаторов, мы с полным правом говорим еще о явлениях плейотропизма. Вместе с тем здесь, однако, совершенно ясно выступают морфогенетические корреляции между отдельными процессами, определяемыми первичными или вторичными морфогенными продуктами. Эти корреляции мы рассматриваем как элементарные формы морфогенетических корреляций, так как в процессе эволюции они всегда претерпевают дальнейшие преобразования и усложнения, в результате которых генетический анализ взаимозависимостей становится практически невозможным. В процессе исторического развития организмов изменения характера мутаций, подобные разобранным, являются лишь базой, на которой строятся видовые различия более крупных таксономических подразделений. В этом случае мы никогда не имеем дела с признаками, определяемыми изменениями единичных генов, а всегда с результатом преобразования очень многих генов, т. е. мы имеем множественную зависимость любого признака и любой фазы его развития. Это может быть пояснено третьей, еще более сложной диаграммой (рис. 39).
Естественно, что система зависимостей, каждое звено которой определяется очень многими факторами, отличается значительно большей устойчивостью, чем разобранные нами ранее выражения простого или многостепенного плейотропизма. Признаки, зависящие в своем развитии от такой сложной системы морфогенетических корреляций (как это характерно для позвоночных), отличаются значительно большей устойчивостью, чем автономные признаки мозаичного организма (например, дрозофилы).
Нормальное развитие всего организма обусловливается при наличии сложной системы корреляций всем генотипом в целом. Роль отдельных генов снижается в связанных частях организма до уровня модификаторов. Система корреляций обеспечивает, таким образом, устойчивость основных черт организации, а возможная здесь без летальных последствий высокая мутабиль-ность отдельных генов гарантирует достаточную пластичность конкретного ее оформления.
В. МОРФОФИЗИОЛОГИЧЕСКИЕ СИСТЕМЫ
По море усложнения системы корреляций, последние теряют свой «генетический» характер, т. е. становятся практически неразложимыми (точнее — их нарушение в результате мутации приводит к летальным последствиям). В таком случае можно говорить о морфофизиологических корреляциях и их системах, как единицах, не поддающихся обычному генетическому анализу. Эти системы корреляций составляются в своей основе из взаимозависимостей в жизненных функциях организма, благодаря которым происходит согласованное течение общефизиологических процессов. В первую очередь это касается клеточного обмена веществ, связанного с образованием морфогенных веществ в развивающемся организме. Затем, это предполагает согласование специфических свойств этих морфогенных веществ с реактивной способностью тканей на различных стадиях развития, т. е. детерминацию и регуляцию формообразования организма (морфогене-тические корреляции), и, наконец, это означает согласование общежизненных функций отдельных частей и органов развивающегося эмбриона, личинки и сформировавшегося организма (собственно физиологические корреляции). Типично физиологические взаимозависимости определяют течение различных жизненно важных процессов, которые в общем характеризуются своей легкой и быстрой обратимостью. Эти взаимозависимости имеют, как правило, регуляторный характер. Их значения мы здесь не рассматриваем, а ограничиваемся морфологическими выражениями зависимостей.
Однако и физиологические зависимости могут обладать своим морфологическим выражением. Это касается некоторых общефизиологических процессов, связанных с ростом и размножением организма, и многих функциональных взаимозависимостей, связанных с побочным морфогенетическим эффектом в виде менее легко обратимых изменений формы и структуры. Это прежде всего взаимное трофическое влияние органов, находящихся в функциональной взаимосвязи. Результатом может быть усиленный рост или ускоренная дифференцировка этих органов («функциональная гипертрофия»).
Это трофическое влияние может частично перейти в детерминирующее влияние, если в результате активации функциональных взаимосвязей происходит развитие тех или иных специфических гистологических или общеморфологических структур (например, структура и форма кости под влиянием деятельности мышц). В этом случае я говорю об эргонтических корреляциях (от греческого эргон — работа). Здесь морфогенетический эффект является, так сказать, побочным результатом существования функциональной взаимозависимости.
Наконец, взаимодействие частей в развивающемся организме может вести уже прямо к формообразовательным процессам. Формообразование является тогда не побочным результатом существования взаимозависимости между двумя или более частями, а основным ее выражением. В этом случае я говорю о морфоге-нетических корреляциях.
Ясно, что между физиологическими, эргонтическими и морфо-генетическими корреляциями имеются все переходы. У взрослого организма формообразовательные процессы сведены к минимуму и взаимосвязь частей выражается в форме физиологических корреляций между легко обратимыми жизненными процессами. У молодого, еще развивающегося организма, лишь вступающего в пору своей дефинитивной жизнедеятельности, эти (физиологические корреляции играют в то же время и роль факторов, обусловливающих развитие многих так называемых функциональных структур, которые дают окончательную отшлифовку развивающимся формам, т. е. выступают в роли эргонтических корреляций. У эмбриона же, у которого почти вся энергия расходуется на формообразовательные процессы, подобные корреляции обусловливают взаимозависимость между этими, характерными для эмбриона, формами жизнедеятельности. Здесь корреляции выступают в роли специфических морфогенетических взаимозависимостей.
Резких граней между этими формами корреляций провести нельзя — они переходят одни в другие, меняя свое выражение по мере развития самого организма. Нетрудно себе также представить и процесс смены форм корреляций во время филогенетического развития организмов, как, например, постепенное замещение некоторых эргонтических корреляций морфогенетиче-скими, по мере «эмбрионализации» самого индивидуального развития.
Вопрос о возникновении новых морфогенетичеоких корреляций не представляет особых трудностей, если к нему подходить с точки зрения морфолога. В процессе эволюции, одновременно с новыми дифференцировками, обязательно устанавливаются и новые взаимозависимости как внутренние факторы, определяющие наследственное осуществление этих дифференцировок (т. е. процессы интеграции являются условием, без которого невозможна и наследственно детерминированная дифференциация). Система корреляций, связывающих развивающийся организм в одно целое, имеет, следовательно, ту же историческую основу, что и сама организация. Поэтому исторически сложившиеся взаимоотношения частей и являются вместе с тем на каждой стадии развития в роли факторов, определяющих дальнейшее течение процессов развития.
Между всеми так или иначе связанными частями обнаруживаются взаимодействия, которые в большей или меньшей мере приобретают детерминирующее значение. Таким образом, типичные морфогенетические корреляции развиваются всегда на базе исторически сложившихся соотношений между частями развивающегося организма. При этом, однако, если установление новых корреляций связано с возникновением любой новой дифферен-цировки, то разрушение старых корреляций неизменно сопровождает редукцию органов, потерявших свое значение.
Г. РАЗВИТИЕ МОРФОГЕНЕТИЧЕСКИХ РЕГУЛЯЦИЙ
Морфогенетические регуляции имеют, конечно, физиологическую, а в конечном счете — именно биохимическую основу. Все же мы их выделяем, поскольку они связаны с защитой нормального формообразования (возникновения собственно физиологических регуляций — огромной области эволюционной физиологии — мы здесь не касаемся). Такой защитой являются уже рассмотренные генетические системы, сформирование которых означает, очевидно, установление более устойчивых форм биохимического равновесия в процессах внутриклеточного обмена и синтеза живого вещества. Целостность таких сбалансированных систем обеспечивается вначале частичной изоляцией отдельных популяций, сложностью самих сочетаний, не допускающих их нарушения (без летальных последствий), диплоидностыо организмов, что гарантирует наличие по меньшей мере, одной полной «нормальной», генетической системы (в двух геномах) в любой почти особи природных популяций, и доминантностью нормальных аллеломорфов. Эти генетические системы защищают до известной степени развитие нормы. Однако не следует думать, что «баланс» представляет особую форму равновесия между «генами». Термины генетиков, например, «генный баланс», доминантные «гены», двойной запас «активности генов» и т. п., служат лишь упрощенными выражениями, удобными для описания сложных явлений. За этими терминами скрываются, однако, очень сложные процессы взаимодействия не «генов», а в первую очередь процессов внутриклеточного обмена веществ, и во вторую очередь — более сложных физиологических и, наконец, морфогенетических процессов. Поэтому все формы «защиты» нормального формообразования, все формы морфогенетических регуляций являются всегда результатом взаимодействия между физиологическими процессами и выражением взаимозависимости частей в корреляционных системах развивающегося организма. Если мы выделяем все же генетические и мор-фофизиологические системы, то это отражает процесс усложнения форм взаимозависимостей, которые в начале своего появления поддаются методам генетического анализа, а в дальнейшем процессе эволюции становятся уже неразложимыми при помощи этих методов (как это было только что показано).
|
|
|
|
|
Дата добавления: 2015-05-06; Просмотров: 677; Нарушение авторских прав?; Мы поможем в написании вашей работы!