КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Митоз, его фазы, биологическое значение
10.
Мембрана (от лат. membrāna «пергамент») — плёнка, выступающая обычно как полупроницаемый разделитель сред (в том числе как оболочка) или как колебательная поверхность.
По своему типу мембраны делятся на пористые и непористые.
В биологии:
- Клеточная мембрана
- Мембрана (экзоскелет членистоногих) — не подвергшиеся склеротизации участки кутикулы членистоногих.
Клеточные и субклеточные биологические мембраны. Типы клеточных мембранных структур (плазматическая мембрана, мембраны клеточного ядра, эндоплазматический ретикулум, аппарат Гольджи, лизосомы, митохондрии). Функции биомембран. История развития представлений о структуре мембран.
Состав биомембран. Липидный состав биологических мембран. Фосфолипиды. Гликолипиды. Стерины. Многообразие функции мембранных липидов. Структура наружных мембран грамотрицательных и грамположительных бактерий.
Белки биологических мембран. Периферические белки, интегральные белки. Углеводы биологических мембран. Методы выделения компонентов мембран.
Методы выделения мембран и их компонентов. Детергенты, используемые в мембранологии. Классификация детергентов. Химическая структура детергентов. Процесс солюбилизации мембранных препаратов детергентами.
Физико-химические методы исследования биомембран. Дифракция рентгеновских лучей. Электронная микроскопия. ЯМР спектроскопия.
Модели биологических мембран. Монослои на границе раздела фаз. Черные мембраны. Эмульсии. Липосомы и их использование в исследовании мембран. Получение липосом. Области применения липосом.
Фазовые переходы липидов в мембране. Переход гель – жидкий кристалл, механизм явления. Методы исследования (дифференциальная сканирующая калориметрия, спектроскопия ядерного магнитного резонанса, флуоресценция). Возможная биологическая роль. Термотропные свойства липидных смесей.
Полиморфизм липидов. Лиотропные жидкие кристаллы, фазовые диаграммы. Структура фаз. Фазовый переход бислой – небислойные структуры. Возможная биологическая роль полиморфизма.
Липид-белковые взаимодействия. Методы изучения липид-белкового взаимодействия. Свойства мембран (текучесть, латеральная диффузия липидов и белков, асимметрия).
Транспортные процессы в мембранах. Транспорт низкомолекулярных веществ. Поры и каналы. Ионофоры. Пассивный транспорт (простая и облегченная диффузия). Активный транспорт (работа ферментов АТФ-аз.) Эндоцитоз, экзоцитоз, их возможная биологическая роль.
Молекулярные основы межклеточной и внутриклеточной коммуникации. Сигнальные молекулы. Пути передачи сигнала внутри клетки. Путь аденозинмонофосфатный и фосфоинозитольный. Вторичные мессенджеры.
Перекисное окисление липидов. Активные формы кислорода: образование и утилизация. Цепной механизм перекисного окисления. Антиоксиданты. Роль перекисного окисления липидов в развитии патологии.
Биогенез биологических мембран. Биосинтез липидов в эукариотических и прокариотических клетках. Внутри- и межмембранный транспорт липидов. Внутриклеточный транспорт мембранных белков.
СПЕЦИАЛИЗА́ЦИЯ, в биологии — приспособление видов и надвидовых таксонов к более узкой, по сравнению с предками, среде обитания. Специализация, как ароморфоз (см. АРОМОРФОЗ)и идиоадаптация, — один из способов достижения биологического прогресса (см. БИОЛОГИЧЕСКИЙ ПРОГРЕСС). Однако, сужение условий существования специализированных видов может привести к их вымиранию при резких изменениях внешних условий. И. И. Шмальгаузен (см. ШМАЛЬГАУЗЕН Иван Иванович) (1939) выделил четыре основные формы специализации: катаморфоз, гипоморфоз, гиперморфоз и теломорфоз.
При катаморфозе (общей дегенерации) организация упрощается вплоть до исчезновения ароморфозов, свойственных предкам. Это направление эволюции чаще всего возникает у эндопаразитов. Например, у цестод (ленточных глистов (см. ЛЕНТОЧНЫЕ ЧЕРВИ)) редуцировались мышечная и пищеварительная системы, нервная система упростилась. Развиты только выделительная и половая. Питательные вещества эти паразиты всасывают из содержимого кишечника или из тканей хозяина всей поверхностью тела. От изменений окружающей среды их защищает хозяин, в котором они живут. Цестоды паразитируют во всех позвоночных животных, а их личинки — во многих беспозвоночных. Класс цестод включает 21 семейство и более 250 родов. Число видов точно не известно, т. к. постоянно описывают новые виды. В этом случае специализация — одна из форм биологического прогресса.
При гипоморфозе организмы утрачивают конечные стадии онтогенеза и переходят к неотении (см. НЕОТЕНИЯ). При этом они теряют и ту часть среды обитания, в которой существовали их взрослые предки. Гиперморфоз — специализация, связанная с увеличением размеров тела вплоть до гигантизма. Наиболее известным примером гиперморфоза служат гигантские динозавры. Гигантизм связан со специализацией по количеству пищи. Огромные травоядные ящеры могли существовать только в теплом влажном климате с богатой растительностью и только они могли обеспечить собой пропитание хищным динозаврам. Слоны сохранились только в тропиках Азии и Африки. Мамонты вымерли с исчезновением холодных приледниковых степей, где торчавшая из под снега сухая трава («сено на корню») обеспечивала их пищей. Последние мамонты погибли 4—5 тысяч лет назад в северной Якутии и на Чукотке, где климат дольше всего был похож на климат ледникового периода.
Наиболее многообразна специализация, называемая теломорфозом. Обычно она связана со специализацией в отношении какого-либо одного источника пищи. Например, муравьед питается только термитами, а колибри — только нектаром цветов. Такая специализация позволяет избегать конкуренции за пищу с другими животными. Однако, как и при гиперморфозе, резкие изменения среды обитания приводят таких животных к быстрому вымиранию
Клеточный цикл — это период существования клетки от момента её образования путем деления материнской клетки до собственного деления.
Содержание
[убрать]
|
[править] Длительность клеточного цикла эукариот
Длительность клеточного цикла у разных клеток варьируется. Быстро размножающиеся клетки взрослых организмов, такие как кроветворные или базальные клетки эпидермиса и тонкой кишки, могут входить в клеточный цикл каждые 12—36 ч. Короткие клеточные циклы (около 30 мин) наблюдаются при быстром дроблении яиц иглокожих, земноводных и других животных. В экспериментальных условиях короткий клеточный цикл (около 20 ч) имеют многие линии клеточных культур. У большинства активно делящихся клеток длительность периода между митозами составляет примерно 10—24 ч.
[править] Фазы клеточного цикла эукариот
Клеточный цикл эукариот состоит из двух периодов:
- Период клеточного роста, называемый «интерфаза», во время которого идет синтез ДНК и белков и осуществляется подготовка к делению клетки.
- Периода клеточного деления, называемый «фаза М» (от слова mitosis — митоз).
Интерфаза состоит из нескольких периодов:
- G1-фазы (от англ. gap — промежуток), или фазы начального роста, во время которой идет синтез мРНК, белков, других клеточных компонентов;
- S-фазы (от англ. synthesis — синтетическая), во время которой идет репликация ДНК клеточного ядра, также происходит удвоение центриолей (если они, конечно, есть).
- G2-фазы, во время которой идет подготовка к митозу.
У дифференцировавшихся клеток, которые более не делятся, в клеточном цикле может отсутствовать G1 фаза. Такие клетки находятся в фазе покоя G0.
Период клеточного деления (фаза М) включает две стадии:
- кариокинез (деление клеточного ядра);
- цитокинез (деление цитоплазмы).
В свою очередь, митоз делится на пять стадий.
Описание клеточного деления базируется на данных световой микроскопии в сочетании с микрокиносъемкой и на результатах световой и электронной микроскопии фиксированных и окрашенных клеток.
[править] Регуляция клеточного цикла
Закономерная последовательность смены периодов клеточного цикла осуществляется при взаимодействии таких белков, как циклин-зависимые киназы и циклины. Клетки, находящиеся в G0 фазе, могут вступать в клеточный цикл при действии на них факторов роста. Разные факторы роста, такие как тромбоцитарный, эпидермальный, фактор роста нервов, связываясь со своими рецепторами, запускают внутриклеточный сигнальный каскад, приводящий в итоге к транскрипции генов циклинов и циклин-зависимых киназ. Циклин-зависимые киназы становятся активными лишь при взаимодействии с соответствующими циклинами. Содержание различных циклинов в клетке меняется на протяжении всего клеточного цикла. Циклин является регуляторной компонентой комплекса циклин-циклин-зависимая киназа. Киназа же является каталитическим компонентом этого комплекса. Киназы не активны без циклинов. На разных стадиях клеточного цикла синтезируются разные циклины. Так, содержание циклина B в ооцитах лягушки достигает максимума к моменту митоза, когда запускается весь каскад реакций фосфорилирования, катализируемых комплексом циклин-В/циклин-зависимая киназа. К окончанию митоза циклин быстро разрушается протеиназами.
[править] Контрольные точки клеточного цикла
Для определения завершения каждой фазы клеточного цикла необходимо наличие в нем контрольных точек. Если клетка «проходит» контрольную точку, то она продолжается «двигаться» по клеточному циклу. Если же какие-либо обстоятельства, например повреждение ДНК, мешают клетке пройти через контрольную точку, которую можно сравнить со своего рода контрольным пунктом, то клетка останавливается и другой фазы клеточного цикла не наступает по крайней мере до тех пор, пока не будут устранены препятствия, не позволявшие клетке пройти через контрольный пункт. Существует как минимум четыре контрольных точки клеточного цикла: точка в G1, где проверяется интактность ДНК, перед вхождением в S-фазу, сверочная точка в S-фазе, в которой проверяется правильность репликации ДНК, сверочная точка в G2, в которой проверяются повреждения, пропущенные при прохождении предыдущих сверочных точек, либо полученные на последующих стадиях клеточного цикла. В G2 фазе детектируется полнота репликации ДНК и клетки, в которых ДНК недореплицирована, не входят в митоз. В контрольной точке сборки веретена деления проверяется, все ли кинетохоры прикреплены к микротрубочкам.
[править] Нарушения клеточного цикла и образование опухолей
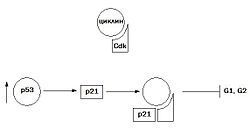

Увеличение синтеза белка p53 ведет к индукции синтеза белка p21 — ингибитора клеточного цикла
Нарушение нормальной регуляции клеточного цикла является причиной появления большинства твердых опухолей. В клеточном цикле, как уже говорилось, прохождение контрольных пунктов его возможно только в случае нормального завершения предыдущих этапов и отсутствия поломок. Для опухолевых клеток характерны изменения компонентов сверочных точек клеточного цикла. При инактивации сверочных точек клеточного цикла наблюдается дисфункция некоторых опухолевых супрессоров и протоонкогенов, в частности p53, pRb, Myc и Ras. Белок p53 является одним из факторов транскрипции, который инициирует синтез белка p21, являющегося ингибитором комплекса CDK-циклин, что приводит к остановке клеточного цикла в G1 и G2 периоде. Таким образом клетка, у которой повреждена ДНК, не вступает в S-фазу. При мутациях, приводящих к потере генов белка p53, или при их изменениях, блокады клеточного цикла не происходит, клетки вступают в митоз, что приводит к появлению мутантных клеток, большая часть из которых нежизнеспособна, другая — дает начало злокачественным клеткам.
Важнейшим компонентом клеточного цикла является митотический (пролиферативный) цикл. Он представляет собой комплекс взаимосвязанных и согласованных явлений во время деления клетки, а также до и после него. Митотический цикл — это совокупность процессов, происходящих в клетке от одного деления до следующего и заканчивающихся образованием двух клеток следующей генерации. Кроме этого, в понятие жизненного цикла входят также период выполнения клеткой своих функций и периоды покоя. В это время дальнейшая клеточная судьба неопределенна: клетка может начать делиться (вступает в митоз) либо начать готовиться к выполнению специфических функций.
|
|
Дата добавления: 2015-04-23; Просмотров: 755; Нарушение авторских прав?; Мы поможем в написании вашей работы!