КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Особенности структуры и биомеханики мышечной ткани
|
|
|
|
Мышцы – это ткани, способные производить механическую работу, и тем самым обеспечивать перемещение человека, движение воздуха в дыхательных путях, движение крови и многие другие, жизненно важные процессы. Различают два основных типа мышц: поперечно-полосатые (прикрепляются к различным частям скелета, поэтому их также называют скелетными мышцами) и гладкие мышцы (миокард, стенки внутренних органов, кровеносных и лимфатических сосудов и др.).
Рассмотрим особенности строения и механизм мышечного сокращения на примере поперечно-полосатой мышцы  . Она состоит из пучков параллельных волокон, каждое из которых имеет диаметр 10 – 100 мкм и длину до 10 – 15 см. Волокно окружено упругой мембраной (сарколеммой) и состоит из множества параллельно располагающихся миофибрилл диаметром 1 – 2 мкм. Миофибрилла в свою очередь состоит из ряда белковых нитей – толстых и тонких, симметричное расположение которых в её поперечном сечении миофибриллы показано на рис. 23. На рис. 24 изображено продольное сечение миофибриллы при её трёх разных длинах. В поляризованном свете миофибрилла имеет периодическую полосатую структуру, так как состоит из оптически изотропных участков (I -зоны) и анизотропных участков (A -зоны). Именно это определило название этой мышцы: поперечно-полосатая. При растяжении покоящейся мышцы в середине А -зоны появляется зона Н меньшей плотности.
. Она состоит из пучков параллельных волокон, каждое из которых имеет диаметр 10 – 100 мкм и длину до 10 – 15 см. Волокно окружено упругой мембраной (сарколеммой) и состоит из множества параллельно располагающихся миофибрилл диаметром 1 – 2 мкм. Миофибрилла в свою очередь состоит из ряда белковых нитей – толстых и тонких, симметричное расположение которых в её поперечном сечении миофибриллы показано на рис. 23. На рис. 24 изображено продольное сечение миофибриллы при её трёх разных длинах. В поляризованном свете миофибрилла имеет периодическую полосатую структуру, так как состоит из оптически изотропных участков (I -зоны) и анизотропных участков (A -зоны). Именно это определило название этой мышцы: поперечно-полосатая. При растяжении покоящейся мышцы в середине А -зоны появляется зона Н меньшей плотности.
Толстые нити миофибрилл образованы миозином, тонкие – актином, они частично перекрываются друг с другом. Каждая толстая нить состоит из 180 – 360 продольно ориентированных молекул миозина, ответственных за анизотропию А -зоны. Изотропные I -зоны образованы относительно неупорядоченными тонкими нитями актина.
|
|
|
Важным элементом миофибриллы является тонкая мембрана (Z -диск), имеющая поры, сквозь которые проходят актиновые нити. Мембрана фиксирует их пространственное расположение и создает условия для их строго определённого смещения при сокращении мышцы.
Участок миофибриллы между двумя Z -дисками называется саркомером. Саркомер – наименьшее образование, которое обладает свойством сократимости. Его структура показана на рис. 25. Каждая миофибрилла окружена специальной мембранной структурой – саркоплазматическим ретикулумом, содержащим ионы кальция (Са++).
 Толстые и тонкие нити взаимодействуют между собой с помощью «головок» миозина, образующих соединительные «мостики».
Толстые и тонкие нити взаимодействуют между собой с помощью «головок» миозина, образующих соединительные «мостики».
Общепринятой моделью мышечного сокращения является модель скользящих нитей, согласно которой сократительный процесс происходит следующим образом. Возбуждаемая нервным импульсом сарколемма электрохимически передает процесс возбуждения на саркоплазматической ретикулум. В результате повышается проницаемость этой мембранной структуры для ионов Са++ и происходит выброс Са++ в цитоплазматическую жидкость (саркоплазму), заполняющую мышечное волокно. Повышение концентрации Са++ с 10–7 до 10–5 моль/л стимулирует циклическую работу миозиновых «мостиков». «Мостик» связывается с актином и тянет его к центру А -зоны, в область расположения миозиновых нитей, перемещая на расстояние 10 – 12 нм. Затем он отщепляется от актина, связывается с ним в другой точке и опять подтягивает в нужную сторону. Непрерывное движение актиновных нитей происходит как результат поочередной работы «мостиков». Частота циклов их движений, по-видимому, регулируется в зависимости от нагрузки на мышцу и может достигать 1000 Гц. «Мостики» обладают АТФ-азной активностью, стимулируют гидролитическое расщепление АТФ и используют освобождающуюся при этом энергию для своей работы.
|
|
|
Возвращение мышцы к исходному состоянию обусловлено с обратными переходами ионов Са++ из саркоплазмы в ретикулум вследствие работы кальциевых насосов. Таким образом, именно ионы Са++ регулируют сократительные процессы в мышцах.
Механическое усилие, развиваемое мышцей при сокращении, зависит от еë поперечного сечения, от начальной длины волокон и ряда других факторов. Сила мышцы, приходящаяся на 1 см2 её поперечного сечения, называется абсолютной мышечной силой. Для человека она изменяется в пределах 50 – 100 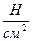 . Сила одних и тех же мышц человека зависит от ряда физиологических условий: возраста, пола, тренированности и т.д.
. Сила одних и тех же мышц человека зависит от ряда физиологических условий: возраста, пола, тренированности и т.д.
Модуль упругости мышцы (модуль Юнга) Е, определённый in vitro, примерно равен 106 Па.
Сократительная активность мышц (пучка мышечных волокон или отдельного волокна) при фиксированной их длине проявляется в развитии ими механического усилия (механического напряжения), а при фиксированной нагрузке – в их укорочении. В соответствии с этим эксперименты с мышцами проводятся при двух основных режимах: изометрическом (когда длина мышцы фиксирована, но может изменяться развиваемое ею механическое усилие) и изотоническом (нагрузка, приложенная к мышце, постоянна и мышца имеет возможность сокращаться).


На рис.26 приведены схемы установок по реализации в эксперименте изометрического (а) и изотонического (б) режимов. Здесь Д F – датчик силы, Д L – датчик изменения длины, М – мышца, Эл – электроды стимуляции, Р – нагрузка, Ф – фиксатор длины.
На рис. 27 представлены результаты этих экспериментов с портняжной мышцей лягушки при 0оС. Видно, что изометрическое усилие развивается очень быстро и достигает максимальной величины примерно через 170 мс после возбуждения мышцы, а начиная с 200-й миллисекунды оно уменьшается, но даже через 900 мс в мышце ещё сохраняется некоторое напряжение, что может быть обусловлено активными физическими и химическими процессами в ней.

 Режим изотонического одиночного сокращения существенно отличается от изометрического режима (см. рис. 27). Укорочение в таком режиме начинается не сразу, а только тогда, когда в мышце развивается достаточное усилие, равное по величине приложенному внешнему усилию. Поэтому сокращение мышцы начинается тем позднее, чем больше внешняя нагрузка. В изотоническом режиме укорочение мышцы сначала растет почти линейно со временем, затем этот рост замедляется и прекращается тем раньше, чем больше нагрузка. Затем наступает расслабление мышцы, которое также завершается тем раньше, чем больше внешняя нагрузка (см. рис.27, б). Скорость укорочения мышцы при фиксированной нагрузке примерно постоянна и зависит от величины этой нагрузки. При минимальной (то есть нулевой) нагрузке скорость укорочения мышцы будет максимальной, а с увеличением нагрузки постепенно уменьшается. Если же сделать нагрузку максимальной Р мах, т.е. равной тому максимальному изометрическому усилию, которое мышца способна развить, то никакого внешнего укорочения её не произойдет.
Режим изотонического одиночного сокращения существенно отличается от изометрического режима (см. рис. 27). Укорочение в таком режиме начинается не сразу, а только тогда, когда в мышце развивается достаточное усилие, равное по величине приложенному внешнему усилию. Поэтому сокращение мышцы начинается тем позднее, чем больше внешняя нагрузка. В изотоническом режиме укорочение мышцы сначала растет почти линейно со временем, затем этот рост замедляется и прекращается тем раньше, чем больше нагрузка. Затем наступает расслабление мышцы, которое также завершается тем раньше, чем больше внешняя нагрузка (см. рис.27, б). Скорость укорочения мышцы при фиксированной нагрузке примерно постоянна и зависит от величины этой нагрузки. При минимальной (то есть нулевой) нагрузке скорость укорочения мышцы будет максимальной, а с увеличением нагрузки постепенно уменьшается. Если же сделать нагрузку максимальной Р мах, т.е. равной тому максимальному изометрическому усилию, которое мышца способна развить, то никакого внешнего укорочения её не произойдет.
|
|
|
На рис. 28. показана зависимость между приложенной нагрузкой Р и установившейся скоростью v укорочения мышцы.
Надо отметить, что оба режима сокращения присущи мышцам не только в искусственных условиях эксперимента. Так, желудочки миокарда в определённую фазу систолы (при закрытых клапанах) работают в изометрическом режиме, а в другую фазу (при открытии полулунных клапанов) – в изотоническом. Жевательные мышцы при сомкнутых челюстях развивают огромное усилие (напряжение) именно в изометрическом режиме. Бицепс плеча человека, наоборот, сокращается обычно в изотоническом режиме.
В изотоническом режиме зависимость между скоростью v мышечного сокращения и приложенной силой F и описывается уравнением Хилла:
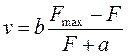 ; (15)
; (15)
Здесь Fmax – максимальное усилие, развиваемое данной мышцей, а – константа, имеющая размерность силы, b – константа, имеющая размерность скорости. Из уравнения (15) следуют выше рассмотренные свойства: при нулевой нагрузке (F=P =0) скорость сокращения мышцы максимальна, а максимальной нагрузке (F мах = P мах) укорочение не происходит (v= 0).
Уравнение Хилла можно получить из анализа механических параметров сокращения и энергетики работы мышцы: энергия Е, освобождающаяся при расщеплении АТФ, превращается частично в механическую работу F D l мышцы, а частично в тепло Q = a D l, которое тоже приближенно пропорционально укорочению D l. Поэтому при изотоническом сокращении мышцы распределение выделившейся энергии можно записать следующим образом:
|
|
|
Е = F D l + a D l = (F + a) D l, (16)
Дифференцируя это уравнение по времени и учитывая, что при изотоническом сокращении сила постоянна (F = const, т.е. не зависит от времени, как и константа а), получим уравнение для мощности, развиваемой мышцей при сокращении:
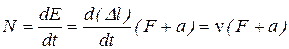 , (17)
, (17)
Вместе с тем экспериментально установлено, что связь между мощностью, развиваемой скелетной мышцей, и ее нагрузкой F достаточно точно описывается следующим соотношением:
N = 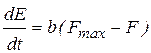 . (18)
. (18)
Сравнив (17) и (18), получим уравнение
n (F + a) = b (Fmax – F); (19)
из которого следует уравнение Хилла (15).
Коэффициент полезного действия (КПД) при сокращении мышцы можно определить как отношение совершённой работы А к затраченной энергии Е:
КПД = 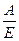 (20) Он может достигать 40- 60% в разных типах мышц.
(20) Он может достигать 40- 60% в разных типах мышц.
Следует отметить, что активные механические свойства мышцы сильно зависят от её пассивного состояния в двигательном аппарате: лишь при определенном пассивном растяжении, обеспечивающем длину саркомера несколько более 2 мкм, мышца работает с максимальной эффективностью.
|
|
|
|
|
Дата добавления: 2015-05-09; Просмотров: 1150; Нарушение авторских прав?; Мы поможем в написании вашей работы!