КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Транспорт факторов вирулентности к мишеням
|
|
|
|
Секреция (экспорт) белков. Схема котрансляционной секреции экзоферментов
Схема экзогенной индукции
Роль фосфотрансферазной системы в катаболической репрессии
Каковы основные приемы получения собственно покоящихся клеток и способы защиты клеточных структур от повреждений
Фосфотрансферазная система состоит из двух неспецифических компонентов: ферментов I и НРг и набора субстрат-специфических белков, связанных с мембраной и обозначенных как ферменты II. Фермент I обеспечивает перенос богатой энергией фосфатной группы от фосфоенолпирувата на гистидиновый остаток фермента НРг, который превращается в фосфо-HPr. Последний является общим донором фосфорильной группы для всех субстратов, переносимых фосфотрансферазной системой. Фосфорилирование же их осуществляется субстрат-специфическими белками из группы ферментов II, которые выполняют также и функции пермеаз. У мутантных бактерий, лишенных фермента I или белка НРг, ферменты II осуществляют облегченную диффузию своих субстратов.

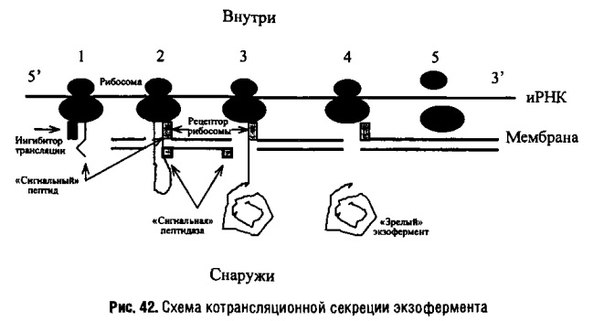
Как известно, синтез белков у бактерий осуществляется на 70s рибосомах, которые также, как и у клеток высших организмов, имеют двоякую локализацию. Большая часть рибосом бактериальных клеток образует полисомы в цитоплазме, около 25% рибосом связано с плазматической мембраной. Такие рибосомы участвуют как в синтезе белков мембраны, так и в синтезе экскретируемых белков. Многие бактериальные клетки получают питательные вещества за счет деградации полимеров около бактериальной поверхности. Для этого бактерии должны выделять гидролизирующие ферменты в окружающую среду. Это они делают намного проще, чем эукариотические клетки: часть их рибосом, локализованных на внутренней (цитоплазматической) поверхности плазматической мембраны, синтезирует белки, которые, подобно секреторным белкам, проходят через мембрану и оказываются вне клетки. Выделенные гидролазы застревают в компонентах муреиновой бактериальной стенки и там функционируют. На других рибосомах, связанных с мембранами, идет синтез белков для построения самой мембраны, подобно тому, что происходит в гранулярном ЭР эукариотических клеток. Так что в этом отношении бактерию можно уподобить вакуоли гранулярного ЭР, вывернутой наизнанку.
|
|
|
Различают два варианта транспорта экзогенных веществ из цитоплазмы: это - секреция и экспорт. Под секрецией понимают транспорт через внутреннюю и внешнюю мембраны, под экспортом - транспорт через цитоплазматическую мембрану в периплазматическое пространство. В настоящее время описаны 4 основных типа секреции протеинов из цитоплазмы микроорганизмов. Для II и IV типа характерно наличие двух отдельных этапов транспорта через внутреннюю и внешнюю мембраны. При этом механизм транспорта через цитоплазматическую мембрану одинаков для обоих типов и предполагает участие комплекса Sec-белков, локализованных во внутренней мембране. Транспорт через внешнюю мембрану у II типа секреции осуществляется без участия дополнительных белков, секретируемый белок формирует пору во внешней мембране благодаря аутокаталитическим процессам, при секреции по IV типу для осуществления этого этапа необходимы дополнительные белки. При транспорте протеинов по I типу секреция происходит в один этап, для ее осуществления необходимы три белка.
Наибольший интерес представляет система секреции III типа, для ее функционирования необходимы 20 белков. Главной особенностью системы III типа является то, что белковые факторы вирулентности секретируются не в окружающую среду, а транслоцируются непосредственно в цитоплазму клеток хозяина, что позволяет рассматривать эту систему как фактор вирулентности. В пользу такой оценки свидетельствует и то, что экспрессия белков системы регулируется в зависимости от условий окружающей среды (контакта с эукариотическими клетками), хотя детали такой регуляции еще не ясны.
|
|
|
Белки системы секреции III типа, выявленные у таксономически далеких микроорганизмов (например, у Pseudomonas aeruginosa и Salmonella typhimurium), характеризуются значительной гомологией, что позволяет с высокой долей вероятности предположить общность их происхождения и последующее горизонтальное распространение среди различных видов грамотрицательных бактерий. Здесь же необходимо отметить, что наличие у таксономически далеких микроорганизмов сходных генов вирулентности является скорее правилом, чем исключением. Гены вирулентности обычно образуют кластеры, получившие название островков патогенности, способные к горизонтальному распространению.В настоящее время нет достаточно убедительных данных, что экспрессия механизмов III типа секреции регулируется посредством двухкомпонентной системы передачи сигнала, однако связь между этими фундаментальными механизмами вирулентности вполне вероятна.
Одним из универсальных регуляторов транскрипции является механизм чувства кворума (quorum sensing). Принцип действия механизма заключается в активации транскрипции специфических генов при достижении порогового уровня связывания белка-активатора транскрипции (LuxR) с низкомолекулярным аутоиндуктором (AI). Описанный механизм опосредует давно известный феномен большей скорости роста культур микроорганизмов при больших величинах посевной дозы. Есть данные об участии этого механизма в регуляции экспрессии детерминант вирулентности P.aeruginosa, следовательно, он также может быть источником мишеней для разработки новых препаратов.
|
|
|
|
|
Дата добавления: 2015-05-07; Просмотров: 1005; Нарушение авторских прав?; Мы поможем в написании вашей работы!