КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Динамика населения популяции и жизненные стратегии
 |
Для природных популяций принято различать:
1) сезонные изменения численности (плотности) популяции;
2) изменения численности (плотности) популяции в различные годы (рис. 4.3).
Рис. 4.3. Типы динамики численности популяции (по С.А. Северцову).
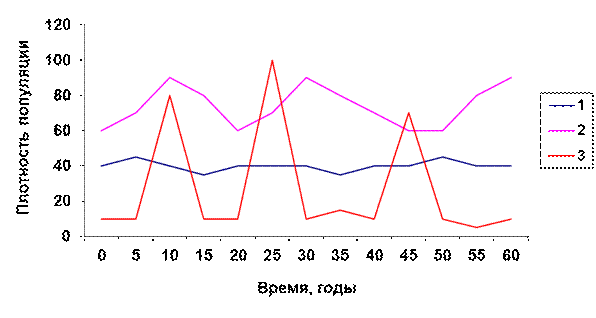
Обозначения к рис. 4.3.
1. Стабильный тип динамики.
2. Лабильный (изменчивый) тип динамики.
3. Эфемерный (взрывной) тип динамики.
Выделенные С.А. Северцовым в начале 40-х годов ХХ века на основе анализа обширного фактического материала основные типы динамики населения популяций птиц и млекопитающих, фактически показали возможность существования ограниченного числа жизненных стратегий (экологических стратегий), позволяющих популяции устойчиво существовать в окружающей среде.
Ещё в 1938 г. выдающийся геоботаник Л.Г. Раменский выделял три основных типа «стратегии жизни», так называемые фитоценотипы растений – виолентов, патиентов и эксплерентов. Позднее Д. Грайм (1979) предложил различать следующие основные стратегии жизни у растений:
конкурентную (конкурентоспособные виды, достигающие высокой плотности популяции в оптимальных местообитаниях; аналогия с виолентами Л.Г. Раменского);
стресс-толерантную (устойчивые к неблагоприятным факторам, но малопродуктивные виды, заселяющие менее благоприятные местообитания; менее корректная аналогия с патиентами);
рудеральную (виды, отличающиеся высоким репродуктивным потенциалом и быстрым ростом, но малой конкурентоспособностью, легко захватывающие нарушенные местообитания; аналогичны эксплерентам Л.Г. Раменского).
Одна из самых известных теоретических концепций стратегий жизни – подразделение организмов на «r – стратегов» и «К – стратегов»:
|
|
|
r-стратегию принято называть «отбором на количество» – теоретически показано (рис. 4.4), что при низкой плотности популяций и высокой доступности ресурсов, в «свободной от конкурентов» среде отбор благоприятствует видам с высоким репродуктивным потенциалом (здесь r соответствует репродуктивному потенциалу в моделях динамики популяции, рис. 4.1 и 4.2), вероятно, r-стратеги соответствуют эксплерентам Л.Г. Раменского и/или рудеральным видам Д. Грайма;
К-стратегию принято называть «отбором на качество» (рис. 4.4); при высокой плотности популяций и ограниченной доступности ресурсов, в «заполненной конкурентами» среде отбор благоприятствует видам с низким потенциалом роста, но высокой способностью конкурировать за ресурсы, повышенной способностью к адаптации и устойчивости (здесь К соответствует «предельной ёмкости среды» в логистической модели динамики популяции, рис. 4.2), вероятно, К-стратеги соответствуют стресс-толерантам Д. Грайма.
Рис. 4.4. Модель r- и К-отбора Мак-Артура (1972, цит. по Ю. Одуму, 1986).
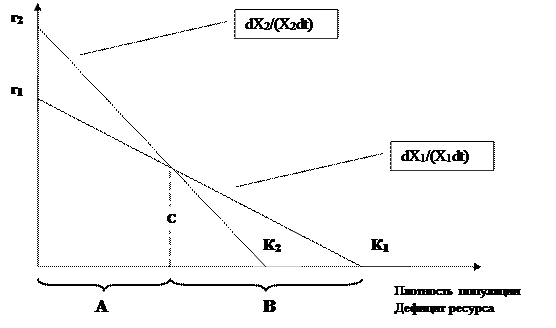 |
Пояснения к рисунку 4.4
Скорости размножения двух аллелей (или видов) изображены в виде функций плотности популяции и ресурса. Когда плотность превышает величину С, а ресурсов мало, Х1 растет быстрее, чем Х2, но Х2 растет быстрее и «победит» в естественном отборе, если плотность ниже величины С. В соответствии с этим А представляет собой область r-отбора, а В – область К-отбора (по Ю. Одуму, 1986).
Ещё одна теоретическая (гипотетическая) модель (рис. 4.5) отражает четыре стратегических пути распределения получаемой организмом (популяцией) энергии между тремя основными видами жизнедеятельности:
1) затратами энергии на размножение;
2) затратами энергии на конкуренцию за общие ресурсы с другими видами (- -);
3) затратами энергии на избегание хищников и другие взаимодействия типа (-; +).
Рис. 4.5. Гипотетическое распределение энергии между тремя основными видами жизнедеятельности (Коуди, 1966, цит. по Ю. Одуму, 1986).
|
|
|
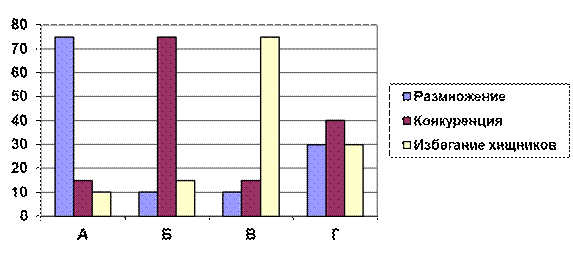
Пояснения к рисунку 4.5
Если ущерб от конкуренции невелик, то большая часть потока энергии может быть направлена на размножение и продуцирование молоди (А); избегание конкуренции или выедания может потребовать больше энергии (Б и В); в случае «Г» энергия распределяется примерно поровну. Случай «А» – обычная ситуация на стадиях колонизации или начальных стадиях сукцессии, когда преобладает r-отбор, тогда как В и Г – характерные ситуации на более зрелых стадиях, когда преобладает К-отбор.
«Естественный отбор, этот бескомпромиссный мастер навязывать функции, требует, чтобы все организмы нашли оптимальное соотношение между расходами на выживание в будущем (… затраты энергии на развитие репродуктивных структур, спаривание, продуцирование семян, яиц, молоди, заботу о потомстве) и расходами на выживание в настоящем (… затраты на компенсацию неблагоприятных экологических факторов, таких как конкуренция за ресурс, хищничество и паразитизм, резкие колебания абиотических факторов» (Ю. Одум, 1986, курсив наш).
На практике отнесение конкретного вида растений или животных к определенному типу «стратегии жизни» наталкивается на многочисленные трудности. Тем не менее, как отмечает И.А. Шилов (1997), «в (предлагаемых различными учеными) экологических стратегиях растений и животных обнаруживается большое сходство, что говорит об общности наиболее фундаментальных экологических основ динамики популяций в разных группах живых организмов».
Авторегуляция плотности населения популяций
Независимая и зависимая от плотности регуляция популяций
Внешние по отношению к популяционной системе экологические факторы, влияющие на динамику плотности населения, принято делить на две группы:
1. Факторы, не зависящие от плотности населения популяции.
Как правило это внешние абиотические экологические факторы (например, температура, влажность, излучения), которые часто (но не всегда) в равной степени воздействуют и на отдельный организм и на группу организмов – не зависят от плотности популяции. Закономерности воздействия таких факторов на отдельный организм, группу организмов или популяцию остаются одинаковыми.
|
|
|
Собственно регулирующими, контролирующими численность популяции, главным образом, путем резкого увеличения смертности, эти факторы могут быть только в экосистемах, находящихся в экстремальных условиях пустыни, тундры и других.
2. Факторы, зависящие от плотности населения популяции.
Это внешние биотические факторы, обусловленные взаимодействием между популяциями различных видов организмов (конкуренция, хищничество, паразитизм и другие), а также механизмы саморегуляции (авторегуляции) плотности населения популяции.
Механизмы авторегуляции плотности населения популяций (по И.А. Шилову, 1997)
У многих видов организмов приближение плотности населения популяции к предельной емкости среды (К) является информационным фактором, сигналом, приводящим в действие запрограммированные в геноме данного вида сложные физиологические и поведенческие реакции, направленные на блокирование размножения (снижение рождаемости и/или увеличение смертности) или усиление эмиграции особей, которые можно рассматривать как механизмы авторегуляции плотности населения популяции.
Классический пример авторегуляции – контроль плотности населения в популяциях грызунов с участием реакции стресса (рис. 4.6).
Рис. 4.6. Схема популяционной авторегуляции плотности населения у грызунов (по С.А. Шиловой, И.А. Шилову, 1977).
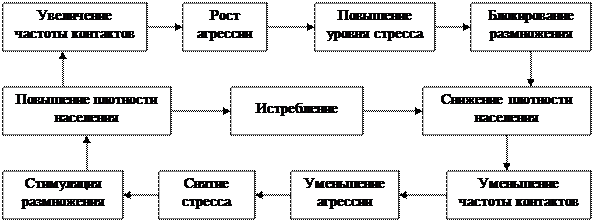
В этом случае социальная напряженность, развивающаяся в популяции при увеличении плотности населения, ведет к повышению уровня стресс-реакции (рис. 4.6.). Последствиями стресса могут быть как увеличение смертности, так и падение рождаемости в популяции.
Во многих случаях универсальным информационным фактором (сигналом), приводящим в действие механизмы авторегуляции плотности населения популяции является так называемое увеличение частоты контактов – увеличение частоты встреч между особями одного вида за определенный промежуток времени.
Можно выделить несколько основных типов авторегуляции плотности населения популяций:
|
|
|
1) регуляция, связанная с конкуренцией за ресурсы – в этом случае недостаток ресурсов (пищи, убежищ, мест и материала для строительства гнезд) служит сигналом (информационным фактором), приводящим к изменению поведения особей (нарушению родительского инстинкта, усилению агрессии по отношению к особям своего вида, проявлению каннибализма);
2) химическая регуляция – зависимая от плотности динамика плодовитости и смертности, связанная с выделением в окружающую среду биологически активных веществ, вызывающих увеличение смертности и/или снижение рождаемости у особей своего вида;
3) регуляция, связанная с изменением поведения – в этом случае сигналом к изменению поведения особей (см. п. 1), приводящего к блокированию размножения или усилению эмиграции, служит увеличение плотности популяции (частоты контактов), что отмечается даже в условиях изобилия ресурсов;
4) регуляция, связанная с территориальным поведением – у территориальных животных, контролирующих определенную территорию, повышение плотности популяции приводит к необходимости тратить основную часть энергии на охрану участка от посягательства других особей своего вида, а не на заботы о продолжении рода;
5) регуляция, связанная с предварительной оценкой плотности населения на данной территории – это поразительный механизм, основанный на особых формах поведения («хоровое» пение лягушек, токование птиц, «рев» у оленей и другие формы «брачных игр», собирающие большое число участников), которые, помимо других функций, позволяют оценить плотность приступающей к размножению популяции. Показано, что плодовитость в таких популяциях зависит от результатов «оценки»;
6) регуляция, связанная со стресс-реакцией – у некоторых высших животных наблюдается сложная иерархическая структура популяций (наличие доминирующих и подчиненных особей). Подчиненные особи испытывают состояние напряжения (стресса), которое усиливается при повышении плотности популяции, что приводит к блокированию размножения (рис. 4.6).
7) регуляция, связанная с эмиграцией (выселением) особей – выселение особей из состава размножающихся группировок – это первая реакция популяции на возрастание плотности населения, позволяющая поддерживать оптимальную плотность без снижения общей численности вида.
«Принцип популяционной авторегуляции плотности исходит из того, что прямая конкуренция за ресурсы может влиять на изменения численности и плотности населения лишь при явной нехватке пищи, убежищ и пр. Такая реакция связана с неизбирательной гибелью особей и может оказывать на популяцию скорее ограничивающее, чем регулирующее действие. Использование информации о плотности населения более эффективно: при этом адаптивная реакция может формироваться раньше, чем наступит реальное истощение ресурсов. По существу, в этом случае прямая конкуренция за ресурсы подменяется конкуренцией за условные объекты: борьба за территорию, полового партнера, положение в иерархии и т. п.» (И.А. Шилов, 1997).
|
|
|
|
|
Дата добавления: 2015-05-10; Просмотров: 1393; Нарушение авторских прав?; Мы поможем в написании вашей работы!