КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Первичная и вторичная зрительная кора. Микро-, макро- и гиперколонки зрительной коры. Формирование зрительных образов. Цветовое зрение
|
|
|
|
Проводниковый отдел зрительного анализатора. Зрительная хиазма. Поле зрения, острота зрения. Бинокулярное зрение. Роль гипоталамуса и четверохолмия в переработке зрительной информации.
Клеточное строение сетчатки: ее тормозные и активационные клетки. Фоторецепторы: палочки и колбочки. Зрительные пигменты: родопсин и иодопсины. Сумеречное и цветовое зрение. Световая и темновая адаптация. Ганглиозные клетки on- и off-типов.
С нейроанатомической точки зрения сетчатка – высокоорганизованная слоистая структура, объединяющая рецепторы и нейроны. Фоторецепторные клетки – палочки и колбочки – расположены в пигментном слое, содержащем эпителиальные клетки, и наиболее удалены от хрусталика. Все волокна зрительного нерва, выходящие из сетчатки, лежат в виде переплетенного пучка, создавая препятствие на пути света при его попадании на рецептор. Такое расположение волокон рецепторов не сказывается на чувствительности последних. Кроме того, в том месте, где они выходят из сетчатки по направлению к мозгу, светочувствительные элементы отсутствуют – это так называемое слепое пятно. Свет, попадающий на сетчатку в области слепого пятна, не воспринимается элементами сетчатки, поэтому остается «дефект» изображения, проецируемого на сетчатку. Палочки и колбочки отличаются как структурно, так и 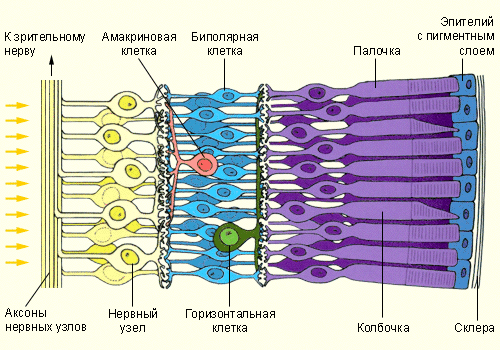
Функционально. Зрительный пигмент родопсин содержится только в палочках. В колбочках находятся другие зрительные пигменты(иодопсин и другие), необходимые для цветового зрения. Палочка гораздо чувствительнее к свету, чем колбочка, но не реагирует на свет с разной длиной волны, т.е. она не цветочувствительна. Зрительные пигменты расположены в наружном сегменте палочек и колбочек. Во внутреннем сегменте находятся ядро и митохондрии, принимающие участие в энергетических процессах при действии света. Зрительный пигмент палочки – родопсин – состоит из белка(опсина) и хромофорной части – ретиналя(альдегид и витамина А). Иодопсин – основной пигмент колбочек- также состоит из опсина и ретиналя. Родопсин и иодопсин имеют разные спектры поглощения: максимум родопсина – 500 нм, иодопсина – 570 нм. Временное ослепление при быстром переходе от темноты к яркому освещению исчезает спустя несколько секунд благодаря процессу световой адаптации. Одним из механизмов световой адаптации является рефлекторное сужение зрачков, другой зависит от концентрации ионов кальция в колбочках. При поглощении света в мембранах фоторецепторов закрываются катионные каналы, что прекращает вхождение ионов натрия и кальция и уменьшает их внутриклеточную концентрацию. Высокая концентрация ионов кальция в темноте подавляет активность гуанилатциклазы — фермента, определяющего образование цГМФ из гуанозинтрифосфата. Вследствие снижения концентрации кальция, обусловленного поглощением света, активность гуанилатциклазы повышается, что ведет к дополнительному синтезу цГМФ. Повышение концентрации этого вещества приводит к открытию катионных каналов, восстановлению тока катионов в клетку и, соответственно, способности колбочек отвечать на световые раздражители как обычно. Низкая концентрация ионов кальция способствует десенситизации колбочек, т. е. уменьшению их чувствительности к свету. Способность различать окружающие предметы исчезает на некоторое время при быстром переходе от яркого света к темноте. Она постепенно восстанавливается в ходе темновой адаптации, обусловленной расширением зрачков и переключением зрительного восприятия с фотопической системы на скотопическую. Темновую адаптацию палочек определяют медленные изменения функциональной активности белков, приводящие к повышению их чувствительности. В механизме темновой адаптации участвуют и горизонтальные клетки, способствующие увеличению центральной части рецептивных полей в условиях низкой освещенности.
|
|
|
|
|
|
Сумеречное зрение характеризуется тем, что в процессе восприятия участвуют как палочки, так и колбочки (специальные рецепторы на сетчатке глаза). То есть если имеется только дневной свет или ночной, то по большей степени, работают только те или другие, а в случае с сумерками, они работают вместе. Поэтому довольно сложно определить точную зависимость светочувствительности. Цветное зрение у человека — это результат работы специальных фоторецепторов, которые помогают различать цвета. Когда свет попадает на сетчатку глаза, как раз именно там и находятся эти нервные клетки. Они бывают двух типов — палочки и колбочки. Палочки работают при ночном зрении и они более светочувствительны, но практически не дают сигналов о цветах, именно поэтому мы видим ночью не так ярко цвета. А вот колбочки наоборот менее светочувствительны, но зато дают возможность более четко различать цвета. Наше цветное зрение — это заслуга этих колбочек. Они бывают 3х видов. Те, которые более чувствительны к зеленому, те, которые лучше воспринимают красный и те, которые воспринимают синий цвета. Наше цветное зрение всего лишь распознает 3 основных цвета(красный, синий, зеленый), а все другие оттенки и цвета получаются при совместной работе этих трех видов колбочек. Цветное зрение — это один из самых изумительных свойств глаза человека, ведь не все животные могут так ярко и четко различать цвета. Нарушение восприятия цвета называется цветовой слепотой или дальтонизмом. Есть три типа нарушений цветового зрения: протанопия – отсутствие чувствительности к красному цвету. Дейтеранопия – к зеленому цвету, тританопия – к синему цвету. Полная цветовая слепота – монохроматия.
В сетчатке описаны 4 типа нейронов: биполярные, горизонтальные, амакриновые и ганглиозные. Палочки и колбочки соединены с биполярными нейронами сетчатки, которые в свою очередь, образуют с ганглиозными клетками синапсы, выделяющие ацетилхолин. Фоторецепторы, соединенные через биполярные нейроны с ганглиозными клетками, образуют их рецептивные поля. После преобразования энергии света в рецепторах сигналы проводятся по двум направлениям: от рецепторов прямо к биполярным, а затем к ганглиозным клеткам сетчатки, и латерально, в боковых направлениях – по слоям горизонтальных клеток(связывающих между собой рецепторы и биполяры) и по слоям амакриновых клеток(связывающих между собой ганглиозные клетки и биполяры). Тва типа тормозных нейронов – горизонтальные и амакриновые клетки – расположены в том же слое, где находятся биполярные нейроны, и функционально связаны как с распространением зрительного возбуждения внутри сетчатки, так и с его ограничением, что определяется структурной организацией внутрисетчаточных связей. Горизонтальные и амакриновые клетки контактируют с биполярными и ганглиозными горизонтальными связями, обеспечивающими латеральное торможение между соседними клеточными элементами сетчатки: горизонтальные – между биполярными, амакриновые – между ганглиозными.
|
|
|
Ганглиозные клетки функционально различны, их разделяют на 3 типа: 1)нейроны, которые возбуждаются светом, падающим на центр рецептивного поля, но затормаживаются, если свет падает на его периферию(«on» - реакция, возникает у нейронов с «on» -центром), 2) нейроны, которые затормаживаются светом в центре рецептивного поля и возбуждаются при действии света на его краях(«off» - реакция, возникает у нейронов с «off» -центром), 3) ней роны, которые отвечают как на включение, так и на выключение света(«on-off»-реакция). Если освещение центральных и переферических участков рецептивного поля происходит одновременно, то для нейронов с «on»-центром преобладает возбуждающий процесс(т.е. увеличение частоты импульсов, связанное с серединой рецептивного поля). У таких нейронов существуют антагонистические отношения между серединой и периферией рецептивного поля. Они выявляются особенно четко при включении светового пятна, освещающего центральную часть рецептивного поля клетки. В этом случае преобладает ответ в виде снижения частоты импульсов, т.е. торможение. И, наоборот, при выключении освещения периферической части рецептивного поля частота импульсации возрастает, т.е. преобладает возбуждение. Нейроны с «off» -центром ведут себя противоположно нейронам с «on» -центром и тоже имеют антагонистические отношения между центром и периферией рецептивного поля. Освещение середины рецептивного поля вызывает уменьшение частоты импульсов, т.е. торможение, а освещение периферической части – возрастание частоты импульсов, т.е возбуждение. При выключении засветки центра рецептивного поля, наоборот, возникает возбуждение, а при выключении освещения периферии – торможение.
|
|
|
Зрительный нерв содержит около 1 млн волокон ганглиозных клеток сетчатки. Зрительные нервы обоих глаз направляются к основанию передней доли гипоталамуса, где пересекаются, образуя хиазму(перекрест). В области основания черепа более полумиллиона волокон зрительного нерва переходят на противоположную сторону. Остальные волокна вместе с перекрещенными аксонами зрительного нерва второго глаза образуют зрительный тракт. После перекреста зрительные пути в виде правого и левого зрительных трактов проходят к следующим структурам мозга: 1) к ядрам верхних бугров четверохолмия(средний мозг) 2)к ядрам наружного(латерального) коленчатого тела(зрительный бугор или таламус)3) к ядрам глазодвигательных нервов4)к супрахиазменным ядрам гипоталамуса. Все аксоны ганглиозных клеток от той половины сетчатки, которая ближе к носу, переходят в области хиазмы на противоположную сторону мозга, проходя в составе зрительного тракта вместе с неперекрещенными волокнами от височной половины сетчатки «своей стороны». Таким образом, после хиазмы вся зрительная информация, относящаяся к левой стороне внешнего мира, воспринимается правой половиной мозга, а относящаяся к правой стороне – левой половиной мозга. Ядра верхних бугров четверохолмия(переднего или верхнего двухолмия) и латерального коленчатого тела являются конечными пунктами 2-х параллельных путей от ганглиозных клеток сетчатки: одна ветвь аксонов ганглиозной клетки идет в верхнее двухолмие, а другая - в латеральное коленчатое тело. Нейроны верхних бугров четверохолмия, на которых проецируются аксоны ганглиозных клеток сетчатки, посылают аксоны к крупному образованию зрительного бугра – подушке, входящей в состав комплекса базальных ганглиев, и к красному ядру, которые учавствуют в регуляции движений. Сигналы об уровне рассеянного света поступают в супрахиазменные ядра гипоталамуса, регулируя уровень бодрствования. Нейроны верхних бугров четверохолмия связаны также с ядрами черепно-мозговых нервов, иннервирующих глазные мышцы. Помимо первичных зрительных проекций от сетчатки глаз нейроны верхних бугров получают сигналы от первичной зрительной коры по петле обратной связи и, кроме того, от нейронов задних бугров четверохолмия – слухового центра среднего мозга. Будучи центром интеграции различных сигналов от зрительной, слуховой и вестибулярной системи (в то же время) от центров координации движения глаз,верхние бугры четверохолмия рефлекторно выполняют важнейшую функцию пространственной ориентации в окружающем постоянно меняющимся мире.
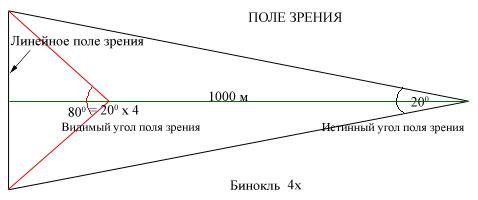
Острота зрения – минимальное различимое глазом угловое расстояние между двумя точками. Острота зрения зависит от общей освещенности окружающих предметов. При дневном свете она максимальна, в сумерках и в темноте острота зрения падает. Наибольшая острота зрения свойствена центральной ямке сетчатки, её желтому пятну. Чем дальше от центральной ямки, тем меньше острота зрения. Острота зрения определяется также организацией нейронной сети сетчатки, а именно – размерами и активностью рецептивных полей. При световой адаптации размер рецептивных полей ганглиозных клетоксетчатки уменьшается, при темной – увеличивается. Динамика размеров рецептивных полей сетчатки отражает приспособление зрительной системы к постоянно изменяющимся условиям освещенности внешней среды, поэтому понижение остроты зрения может быть диагностическим признаком нарушений как в оптическом, так и в нернвном отделах зрительной системы.
Поле зрения – пространство, видимое глазом при фиксации взора в одной точке. Каждый глаз имеет соответственное монокулярное поле зрения. Когда для рассматривания внешнего мира используют оба глаза, то общее поле зрения расширяется. Оно называется бинокулярным полем зрения. В нем имеется центральная область – зона перекрытия монокулярных полей – и две периферические области – с обеих сторон от центральной – каждая для одного глаза. В полях зрения существует такая область, которая вообще не воспринимается. Это так называемое «слепое пятно». С точки зрения структурно-функциональных особенностей сетчатки слепое пятно соответствует месту выхода зрительного нерва из глазного яблока. В этом месте нет никаких рецепторов.
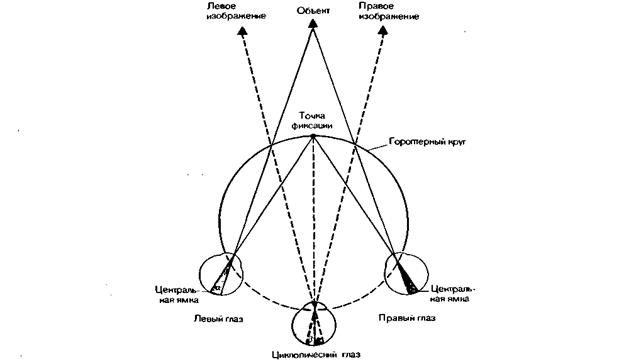
Бинокулярное зрение- зрение с помощью обоих глаз. Оно создается за счет объединения двух монокулярных изображений объектов, усиливая впечатление глубины зрительного пространства. Бинокулярное зрение обеспечивает получение единого сочетанного зрительного образа. Оно осуществляется в результате совместной деятельности сенсорных и моторных систем обоих глаз, обеспечивающих одновременное направление зрительной оси каждого глаза на объект фиксации взора. Построение единого зрительного образа возможно при условии одновременного раздражения центральных ямок или симметрично расположенных по отношению к ним точек обеих сетчаток, так называемых корреспондирующих точек.
Первичная зрительная кора. Первичная зрительная кора лежит в области шпорной борозды, распространяясь вперед от полюса затылка по медиальной стороне каждой затылочной доли. Эта область является конечной станцией для прямых зрительных сигналов от глаз. Сигналы от области макулы сетчатки заканчиваются в районе полюса затылка, тогда как сигналы от периферических частей сетчатки заканчиваются внутри или около концентрических полукругов, расположенных впереди от полюса вдоль шпорной борозды на медиальной стороне затылочной доли. Верхняя часть сетчатки представлена сверху, а нижняя — снизу. На рисунке видно, что макула представлена особенно большим участком первичной зрительной коры. Именно к этой области приходят сигналы от центральной ямки сетчатки, ответственной за наибольшую степень остроты зрения. В общей зоне представительства сетчатки в первичной зрительной коре центральная ямка занимает пространство в несколько сотен раз больше, чем периферические части сетчатки. Первичную зрительную кору называют также зрительной областью I. Другое ее название — стриарная кора, поскольку эта область внешне имеет выраженную полосатую исчерченность. Вторичные зрительные области. Вторичные зрительные области, называемые также зрительными ассоциативными зонами, лежат латерально, впереди, выше и ниже по отношению к первичной зрительной коре. Кроме того, большая часть этих областей накладывается на латеральные поверхности затылочной и теменной коры в виде складок, направленных наружу. В эти области проводятся вторичные зрительные сигналы для анализа их значения. Например, со всех сторон вокруг первичной зрительной коры находится поле Бродмана 18, куда направляются практически все сигналы от первичной зрительной коры. Поле Бродмана 18 называют зрительной областью II, или просто V2. Другие, более отдаленные вторичные зрительные области имеют специфические обозначения V3, V4 и т.д. (более дюжины областей). Значение всех этих областей заключается в постепенном выявлении и анализе различных аспектов зрительного образа.
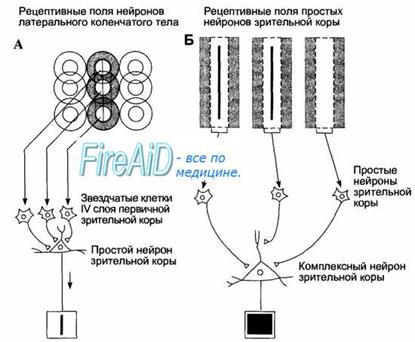
Пирамиды третьего слоя коры собираются в объединения, отграниченные друг от друга анатомически – в виде колонок близко расположенных нейронов. Эти объединения также формируются и по функциональному типу: нейроны, входящие в колонку, обрабатывают одну и ту же информацию, приходящую на колонку по одному афферентному волокну из таламуса. Кроме того, они управляются общей горизонтальной клеткой первого слоя коры, как единое образование. В колонках и формируются представления о событиях, составляющие модель мира.
В зависимости от степени сложности обрабатываемой информации в зрительной коре выделено три типа колонок. Микроколонки реагируют на отдельные градиенты выделяемого признака, например на ту или другую ориентацию стимула (горизонтальную, вертикальную или другую). Макроколонки объединяют микроколонки, выделяющие один общий признак (например, ориентацию), но реагирующие на разные значения его градиента (разные наклоны — от 0 до 180°). Гиперколонка, или модуль, представляет локальный участок зрительного поля и отвечает на все стимулы, попадающие на него. Модуль — вертикально организованный участок коры, выполняющий обработку самых разнообразных характеристик стимула (ориентации, цвета, глазодоминантности и др.). Мо дуль собирается из макроколонок, каждая из которых реагирует на свой признак объекта в локальном участке зрительного поля (рис. 9). Членение коры на мелкие вертикальные подразделения не ограничивается зрительной корой. Оно присутствует и в других областях коры (в теменной, префронтальной, моторной коре и др.).
В коре существует не только вертикальная (колончатая) упорядоченность размещения нейронов, но и горизонтальная (послойная). Нейроны в колонке объединяются по общему признаку. А слои объединяют нейроны, выделяющие разные признаки, ноодинакового уровня сложности. Нейроны-детекторы, реагирующие на более сложные признаки, локализованы в верхних слоях.
Таким образом, колончатая и слоистая организации нейронов коры свидетельствуют, что обработка информации о признаках объекта, таких, как форма, движение, цвет, протекает в параллельных нейронных каналах. Вместе с тем изучение детекторных свойств нейронов показывает, что принцип дивергенции путей обработки информации по многим параллельным каналам должен быть дополнен принципом конвергенции в виде иерархически организованных нейронных сетей. Чем сложнее информация, тем более сложная структура иерархически организованной нейронной сети требуется для ее обработки.
Цветовое зрение, цветное зрение, цветовосприятие, способность глаза человека и многих видов животных с дневной активностью различать цвета, т. е. ощущать отличия в спектральном составе видимых излучений и в окраске предметов. Видимая часть спектра включает излучения с разной длиной волны, воспринимаемые глазом в виде различных цветов. Ц. з. обусловлено совместной работой нескольких светоприёмников, т. е. фоторецепторов сетчатки разных типов, отличающихся спектральной чувствительностью. Фоторецепторы преобразуют энергию излучения в физиологическое возбуждение, которое воспринимается нервной системой как различные цвета, т.к. излучения возбуждают приёмники в неодинаковой степени. Спектральная чувствительность фоторецепторов разного типа различна и определяется спектром поглощения зрительных пигментов. Каждый светоприёмник в отдельности не способен различать цвета: все излучения для него отличаются лишь одним параметром — видимой яркостью, или светлотой, т.к. свет любого спектрального состава оказывает качественно одинаковое физиологическое воздействие на каждый из фотопигментов. В связи с этим любые излучения при определённом соотношении их интенсивностей могут быть полностью неразличимы друг от друга одним приёмником. Если в сетчатке есть несколько приёмников, то условия равенства для каждого из них будут различными. Поэтому для сочетания нескольких приёмников многие излучения не могут быть уравнены никаким подбором их интенсивностей.
Цвет воспринимается зрительной системой (сетчаткой) в условиях достаточной освещённости, необходимой для работы рецепторов-колбочек. Светочувствительные колбочки в ответ на попадание фотонов в глаз стимулируют нервные импульсы, поступающие в мозг, причем импульсы из правого глаза идут в левое полушарие мозга, и наоборот.
Восприятие цвета у млекопитающих происходит через фоточувствительные рецепторы палочки и колбочки, содержащие пигменты с различной спектральной чувствительностью. У большинства приматов, близких к людям, обнаружено несколько типов фоточувствительных пигментов.
На сегодня известны и подробно описаны родопсин (содержащийся в палочках), эритролаб и хлоролаб содержащиеся в колбочках. Трёхкомпонентная гипотеза зрения предполагает наличие в сетчатке глаза трёх типов (по спектрам фоточувствительности) колбочек.
Из предположений трёхкомпонентной гипотезы зрения считается, что видение цвета у большинства приматов — трёхкомпонентное «трихроматики». Остальные приматы и часть млекопитающих, с точки зрения трёхкомпонентной теории цветовосприятия — дихроматики. Принято считать, что у части животных вообще отсутствует цветное зрение.
+еще о гиперколонках.
Гиперколонки или модули первичной зрительной коры
Было сделано предположение, что один полный набор ориентационных колонок, набор цветодетектирующих капель и набор глазодоминантных колонок левого и правого глаза образуют физиологическую единицу первичной зрительной коры. Такая структура получила название гиперколонки или модуля, связанного с определенной частью сетчатки. Рецептивные поля ориентациооно-избирательных клеток в гиперколонке различаются в размерах, но накладываются друг на друга практически в одном и том же месте сетчатки. Вместе они образуют агрегатное рецептивное поле гиперколонки.
|
|
|
|
|
Дата добавления: 2015-05-26; Просмотров: 4755; Нарушение авторских прав?; Мы поможем в написании вашей работы!