КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Эвристическая модель сопряженной эволюции сообщества и биоты
|
|
|
|
Ценотическая регуляция филогенеза. Эта модель, как отмечалось, является дальнейшим развитием идей В. В. Жерихина. В его работах главное внимание обращено на регулирующее воздействие структуры адаптивных зон и экологических ниш сформированного сообщества на процессы филогенеза, входящих в его состав — ценофильных таксонов (Жерихин, Расницын, 1980; Жерихин, 1987).
Такую ценотически регулируемую эволюцию Жерихин вслед за В. А. Красиловым (1969, 1989) называет когерентной. Природу срединномелового биоценотического кризиса, обнаруженного и проанализированного Жерихиным (1978—1987), он объясняет снятием ценотических ограничений филогенеза вследствие разрушения структуры адаптивных зон и экологических ниш биоценоза, и переходом эволюции таксонов в некогерентную фазу. Снятие ограничений обеспечивает быструю в геологическом масштабе времени перестройку сообщества растений, насекомых (Жерихин, 1978, 1980) и ряда других групп животных суши (Жерихин, 1979). Аналогичное событие в ранней—средней юре, но избирательно коснувшееся только сообщества наземных тетрапод, которому посвящена первая часть статьи, мы склонны объяснить так же. Именно события подобного рода, хотя и косвенно, убеждают в существовании и эффективности ценотической регуляции филогенеза в сформированном сообществе.
Однако, если причина кризиса внутренняя, то, указав ближайшую — непосредственную его причину, остается открытым вопрос о процедуре ее исторической подготовки. Вариант ответа на этот вопрос, в частности, предлагает рассматриваемая ниже модель.
Наша задача обратить внимание на противоположный процесс: регуляцию эволюции сообщества процессами филогенеза таксонов. Такое дополнение представлений Жерихина позволяет предложить модель саморазвития сообщества (филоценогенеза) и таксономического разнообразия биоты (филогенеза) по принципу обратной связи. При этом противоречивые отношения в системе сообщество—биота оказываются в конечном счете главной движущей силой их сопряженного развития.
|
|
|
Материал. В основу оригинальной части нашего построения легли распространенные в палеонтологической летописи феномены динамики таксономического разнообразия разных, но экологически сопряженных групп организмов, составляющих одно широко понимаемое сообщество в процессе его исторического развития. Типичная последовательность этих феноменов схематически представлена в левой части рис. 4. История сообщества начинается явным преобладанием относительно малоспециализированных форм, разнообразие которых резко сокращается по мере прогрессивной специализации его членов. Последнее сопровождается дивергенцией и ростом таксономического разнообразия. Затем разнообразие специализированных форм начинает сокращаться без видимых причин, т. е. в отсутствие явно превосходящего по экологической валентности конкурента, и на этом фоне начинает вновь расти разнообразие малоспециализированных форм. Предположение о конкурентном вытеснении первых вторыми является очевидным экологическим нонсенсом. И наконец, в довершение всего возникает новая формация специализированных таксонов, на фоне роста разнообразия которых продолжает сокращаться разнообразие специалистов прежней формации и вновь уменьшается разнообразие малоспециализированных форм. При этом, по крайней мере в начале процесса, первые должны уступать по уровню специализации вторым и потому не могли их вытеснять конкурентно. Это дополнительно подтверждается тем, что сокращение разнообразия специалистов первой формации в подобных случаях начинается задолго до того, как специалисты второй формации становятся многочисленны и разнообразны.
|
|
|
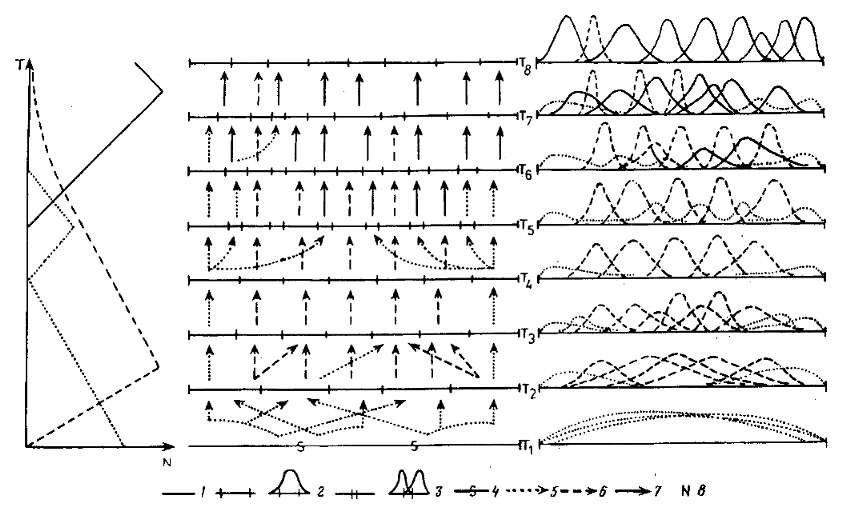
Рис. 4. Модель сопряженной эволюции сообщества, его таксономического и экологического разнообразия: 1 — пространство ресурсов сообщества, 2 — адаптивная зона, 3 — интерзональная экологическая лакуна, 4 — нечетко выраженная граница адаптивных зон, 5 — слабо специализированный таксой, 6—высоко специализированный таксон первой формации, 7—высоко специализированный таксон второй формации, 8— мера таксономического разнообразия
Вся картина в целом начинается экологически очевидным феноменом конкурентного вытеснения малоспециализированных форм более специализированными, но далее приобретает характер процесса, не имеющего тривиальной экологической интерпретации. Приведем два подобных примера из нашей практики.
1 -й пример. На рис. 5, а показана динамика таксономического разнообразия плотоядных зверей Северной Америки. Часть результатов ее анализа была опубликована раньше (Раутиан, 1980). По мере роста разнообразия специализированных хищников палеогена (в крупном размерном классе: Arctocyonia и Crcodonta; в среднем: Miacidae; в мелком — древние группы насекомоядных с относительно низким уровнем специализации: Pantolestidae, Adapisoricidae, Solenodontia, Leptictida) резко сокращается разнообразие самых низко специализированных плотоядных — Didelphoidea, доминировавших в мелу. Их разнообразие вновь подскакивает в олигоцене на фоне сокращения разнообразия перечисленных специалистов. Вслед за этим доминирующими специализированными хищниками становятся Feloidea и Canoidea, занимающие постепенно все размерные классы хищников, а специализированными насекомоядными, так или иначе ушедшими от прямой конкуренции с хищниками (в отличие от более древних вытесненных групп), становятся Erinaceidae, Talpidae и Soricidae. Они составляют вторую формацию специалистов. Падение разнообразия специалистов первой формации начинается задолго до того, когда специалисты второй формации становятся многочисленны и разнообразны. Именно это позволяет легко различить специалистов разных формаций. Таким образом, смену специалистов разных формаций не удается объяснить посредством конкурентного вытеснения. Далее, на фоне роста разнообразия специалистов второй формации, достигшего максимума в миоцене, вымирают сумчатые (Didelphidae). Однако уже в плейстоцене на фоне снижения разнообразия специалистов опоссум (Didelphis) проникает из Южной Америки в Северную, где этот ценофоб в наше время в условиях хозяйственной деятельности человека расширяет географическую область обитания и экологическую сферу своей активности, являясь одновременно вредителем и не очень ценным пушным зверьком.
|
|
|
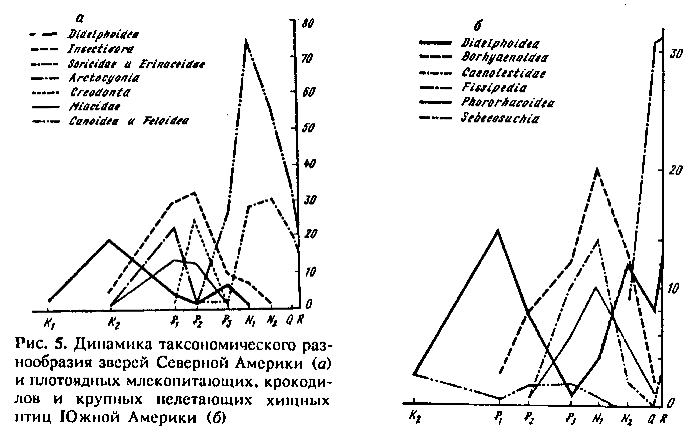
2-й пример, кратко изложенный нами раньше (Каландадзе, Раутиан, 1979), касается плотоядных Южной Америки (рис. 5, б). В мелу и палеогене здесь тоже доминировали Didelphoidea. Позже их сменили в качестве экологических доминантов специализированные хищные (Borhyaenoidea) и насекомоядные (Caenolestidae) сумчатые. Ввиду продолжительного для этого материка дефицита крупных хищных зверей (Каландадзе, Раутиан, 1989) к ним добавились крупные наземные хищные крокодилы (Sebecidae) и многочисленные крупные бегающие хищные птицы (Phororhacoidea). Впечатляющий вторичный рост разнообразия Didelphoidea объясняется прежде всего экологическим кризисом, связанным, по терминологии В. В. Жери-хина, с коллизией биот: внедрением в длительно (с мела, см. рис. 1) изолированную страну (Каландадзе, Раутиан, 1982—1989; Раутиан, 1982) фауны из Северной Америки, обладающей значительно большей экологической валентностью. Это свидетельствует о росте разнообразия малоспециализированных форм в эпоху кризиса. Однако рост разнообразия Didelphoidea, падение разнообразия специализированных хищников и насекомоядов, а также вымирание наземных крокодилов началось задолго до внедрения в Южную Америку североамериканских зверей.
Постановка задачи. Таким образом, предлагаемая модель должна разрешить по крайней мере три экологических парадокса: 1) падение разнообразия специализированных таксонов в отсутствие конкурентов, превосходящих их по экологической валентности; 2) вторичный рост разнообразия малоспециализированных таксонов на фоне высокого разнообразия специалистов; 3) смену специализированных таксонов первой формации таковыми второй формации при условии, что по крайней мере в начале процесса вторые должны уступать по уровню специализации первым и потому не могли их вытеснять конкурентно.
|
|
|
К этому добавляется возникшая еще при чтении первой части статьи проблема исторической подготовки крупного экологического кризиса, которую в первом приближении удается обсудить в связи с решением экологических парадоксов.
Типичная последовательность стадии эволюции сообщества схематически показана на рис. 4. Левая часть его уже рассматривалась выше. В средней части внимание обращено на разделение пространства ресурсов между адаптивными зонами разных таксонов, на характер границ этих зон и преемственную связь (стрелки) обладателей зон на предшествующей и последующей стадиях эволюции сообщества. В правой части метод реализованной экологической ниши (Одум, 1975) применен для характеристики адаптивных зон и отражает величину конкурентного давления (фактического или потенциального), осуществляемого таксонами — обладателями этих зон в разных областях пространства ресурсов.
Филогенетическая и экологическая специализация. Разрушение структуры сообщества, которое в соответствии с концепцией Жерихина, ведет к резкому сокращению таксономического разнообразия в эпоху кризиса и уменьшению общей «суммы жизни», как ее понимал В. И. Вернадский (I960), в свою очередь, ведет к резкому снижению роли конкуренции и прочих факторов, зависящих от плотности, что является важным фактором замедления процесса вымирания в разгар кризиса. Относительно немногие уцелевшие таксоны ведут себя как экологически относительно неспециализированные независимо от их предшествующей филогенетической специализации (Вахрушев, 1988), которая была ориентирована на структуру прежнего сообщества, разрушенного в ходе кризиса.
Различие экологической и филогенетической специализации можно проиллюстрировать примером. На почвах, далеких от педокли-макса, в результате лесопосадок вырастает дубовое криволесье. Здесь дуб — эдификатор климакса — помещен в положение ценофоба. Иными словами, филогенетическая специализация обозначает филогенетическую адаптацию особей данного вида или таксона более высокого ранга. Экологическая специализация обозначает эффективность использования филогенетической специализации в организации конкретной жизнедеятельности. Значительная экологическая специализация в отсутствие филогенетической является структурно-функциональным и экологическим нонсенсом. Всякая экологическая специализация должна иметь морфофункциональный фундамент в организации организма и/или популяции. Очевидно, что в ходе экологического кризиса эффективность филогенетической специализации ценофилов резко падает, а вместе с ней неизбежно падает и экологическая специализация.
Преимущество низкой специализации. Сохранившиеся в ходе кризиса таксоны являются, по существу, «обломками» разрушенного сообщества и экологически широко распространившимися ценофобами. Высокий уровень филогенетической специализации бывших ценофилов работает против своих обладателей, затрудняя смену направления специализации и повышая тем самым вероятность вымирания в ходе кризиса. Сами же черты этой специализации выступают в роли инадаптивного груза (Раутиан, 1988). Поэтому в период кризиса на фоне общего сокращения таксономического разнообразия растет доля низкоспециализировадных форм, в том числе филогенетических реликтов (Жерихин, 1978, 1979, 1987; Каландадзе, Раутиан, 1983).
Экологическое «притяжение». Относительно низкий уровень адаптации большинства таксонов в очень непостоянной биотической среде вызывает высокий уровень элиминиции, который в сочетании с низким уровнем конкуренции создает агрессивную среду обитания (Шмальгаузен, 1968). В этих условиях преимущество получают формы, способные использовать своих экологических соседей в качестве партнеров по совместному выживанию. Действительно, не следует делать адаптивную ставку на ресурс, который (в силу особенностей предшествующей филогенетической специализации) сосед употребляет немного эффективнее или даже если он просто более плодовит и/или многочислен. Естественно попытаться эксплуатировать результаты жизнедеятельности соседа и т. п. Такое экологическое «притяжение» может получить широкое паспростра-нение только в отсутствие жесткой конкуренции и острого дефицита почти на все жизненные ресурсы. Последнее будет характерно для несформированного сообщества, поскольку эффективная эксплуатация даже небогатых ресурсов, в частности их достаточно полная утилизация — признак специализации членов сформированного сообщества. В период кризиса главным дефицитом является адаптивность — уровень специализации. Таким образом, осуществляется первый шаг на пути к коэволюции, ценофилии и прогрессивной специализации таксонов, приступающих к формированию нового сообщества.
1-я фаза филоценогенеза (см. рис. 4, Т1, Т2). Первый результат движения в этом направлении выражается в начале дифференци-ровки пока еще нечетко отграниченных друг от друга адаптивных зон. Это начало процесса роста общего таксономического разнообразия и постепенного снижения роли малоспециализированных таксонов в сообществе.
Коадаптация таксонов дает им взаимные преимущества перед стратегией адаптации ценофобов, не ориентированной на сколько-нибудь определенную (стабильную) биотическую среду. В результате начинаются процессы:
1) постепенного оттеснения ценофобов, широко распространившихся во время кризиса, на периферию зарождающегося сообщества, т. е. в географическое и экологическое пространство, не охваченное процессами филоценогенеза — экотоны;
2) медленного наступления сообщества, охват все большего пространства процессами филоценогенеза и сокращения площади экотонов. Эти процессы — первый сигнал нарастаюшей «суммы жизни» и проявления, еще вялого, конкурентных отношений, ускоряющих и усиливающих специализацию ценофилов и дробление пространства ресурсов на адаптивные зоны.
2-я фаза филоценогенеза (см. рис. 4, Т3). Чем дальше заходит процесс специализации, тем более четкими становятся границы адаптивных зон, тем труднее преодолеваются эти границы и в экологическом и особенно в эволюционном смысле. Сообщество постепенно приобретает черты замкнутости по отношению к иммигрантам. Усиление конкуренции и роли других факторов, зависящих от плотности, постепенно заменяют отношения экологического «протяжения», характерные для несформированного и ненасыщенного сообщества, отношениями конкурентного «отталкивания». При этом биотическая среда, постепенно, по мере роста полноты утилизации ресурсов членами сообщества, приобретает депрессивные черты (Шмальгаузен, 1968).
Предел дроблению адаптивных зон и экологических ниш, как показал В. В. Жерихин, устанавливается емкостью каждой из них, которая не может иметь ресурсов меньше того, что может обеспечить устойчивое существование в ней соответствующего таксона на протяжении филогенетически значимых отрезков времени. Однако прогрессивная специализация таксона — владельца адаптивной зоны — стремится сократить эту емкость за счет роста эффективности эксплуатации ресурсов, стимулируя тем самым дробление адаптивных зон. Поскольку при любой специализации устремить ресурсы к нулю невозможно, то насыщение числа адаптивных зон и экологических ниш неизбежно наступает, хотя это число и не является строго фиксированным.
В конечном счете уже в результате внутризональной конкуренции число адаптивных зон и экологических ниш стремится к числу занимающих их таксонов. Следовательно, ограничение числа адаптивных зон и экологических ниш ограничивает и таксономическое разнообразие биоты сообщества.
3-я фаза филоценогенеза (см. рис. 4, Т4). Интенсификация конкуренции и прогрессивной специализации в освоении биотической среды стимулируют, в порядке положительной обратной связи, дальнейшую специализацию в ранее избранном направлении и ее скорость (Раутиан, 1988). Это, в свою очередь, стимулирует выделение инадаптивных направлений специализации, ведущих к относительно быстрому тактическому успеху.
Возникновение плотной упаковки адаптивных зон практически исключает филогенетически значимые переходы из одной зоны в другую и вселение иммигрантов в данное сообщество. Последнее как массовое явление оказывается возможным только в случае коллизии биот. Такое событие, как отмечалось, произошло в плио-плейстоцене с фауной Южной Америки, ввергнутой в экологический кризис иммигрантами из Северной Америки.
4-я фаза филоценогенеза (см. рис. 4, Т5). Плотная упаковка и жесткое разделение адаптивных зон прекращает активную конкуренцию между владельцами разных зон. Насыщение адаптивных зон знаменует максимум таксономического разнообразия биоты сообщества и перенос напряженных конкурентных отношений внутрь зон. Начинается процесс вытеснения менее специализированных (менее конкурентоспособных) и инадаптивных таксонов с критическим грузом инадаптивности. Одной из наиболее страдающих на этом этапе экологических групп оказываются ценофилы — генералисты, не способные по всей своей широкой адаптивной зоне конкурентно противостоять узким специалистам.
Понижается таксономическое разнообразие и повышается удельный вес узкоспециализированных форм, который в этой фазе филоценогенеза достигают максимума. В условиях плотной упаковки и насыщенности адаптивных зон даже очень небольшое преимущество в специализации (трудно обнаружимое на палеонтологическом материале) может оказаться решающим в сохранении или вымирании формы. Таким образом, получает объяснение первый экологический парадокс: снижение разнообразия специалистов «без видимых причин».
Более специализированная часть вытесняемых таксонов обычно в конечном счете вымирает. В момент наиболее жесткой конкуренции и максимума таксономического разнообразия в эту категорию могут попасть формы, способные жить в данном сообществе не только до этого момента, но и после него, когда в результате вымирания и вытеснения напряженность конкуренции вторично снижается. Примерами могут служить миоценовое вымирание сумчатых и плейстоценовое вымирание лошадей в Северной Америке. Уже отмечалось, что опоссум заселил Северную Америку в плейстоцене, а завезенные туда в историческое время лошади широко расселились в виде мустангов.
Менее специализированная часть вытесняемых таксонов может пополнить ряды ценофобов, изменив стратегию адаптации. Таким образом, разнообразие ценофобов может пополниться за счет неудачливых ценофилов.
Наиболее специализированные таксоны-вытеснители, как правило, не способны обеспечить такое же конкурентное давление, как вытесняемые ими формы по всему пространству ресурсов экологической ниши последних. Об этом косвенно свидетельствует второй экологический парадокс: близкое хронологическое совпадение понижения разнообразия специалистов (в результате внутризональной конкуренции) с началом вторичного роста разнообразия малоспециализированных форм в сообществе, рекрутируемых, очевидно, из экотонов. Именно такое внедрение малоспециализированных форм в сообщество создает ложное впечатление вытеснения ими специалистов.
Адаптивный компромисс. Неспособность вытеснителей полностью скомпенсировать уровень конкурентного давления, который обеспечивали вытесненные ими формы, является почти необходимым следствием адаптивного компромисса (Расницын, 1984—1987) и вытекает из фундаментального запрета на параллельную (одновременную) оптимизацию многих параметров любой системы (Эшби, 1959, 1962). Это утверждение, образно выражаясь, означает, что учиться летать как птица и одновременно бегать как лошадь еще труднее, чем делать то и другое, уже умея. Прогрессивная специализация всегда связана с оптимизацией очень немногих — экзосоматических функциональных систем организма, образующих основное звено его развития (Руженцев, I960). Все же прочие — эндосоматические функциональные системы — эволюируют в координационном порядке (Раутиан, 1988). Следовательно, параллельно со специализацией должна нарастать стенобионтность. Исключение составляют лишь случаи подъема на новый, более высокий уровень организации— событие почти невероятное в рамках сформированного сообщества (Жерихин, 1978, 1979, 1987; Расницын, 1987). Поскольку вытесняемые формы менее специализированы, они с высокой вероятностью окажутся хотя в чем-то эврибионтнее своих вытеснителей.
Фундаментальная и реализованная ниши. Такой взгляд хорошо согласуется с современной экологической парадигмой. В работе Ю. Одума (1975) показаны соотношения фундаментальной и реализованной ниш при разных условиях конкуренции. Чтобы придать им интересующий нас ракурс, достаточно вспомнить, что в процессе филогенеза фундаментальная ниша будет стремиться к реализованной. В нашем случае интенсивная межгрупповая конкуренция приводит к сужению фундаментальной ниши.
Экологические лакуны. Возникающие рассмотренным выше образом зоны ослабленного конкурентного давления мы назвали «экологическими лакунами» (Каландадзе, Раутиан, 1984). Емкость лакуны, по началу во всяком случае, не позволяет поместить в нее мало-мальски специализированный таксой, для которого она фактически не существует. Именно по этой причине из нее оказался вытесненным ее прежний хозяин. Однако для малоспециализированного эврибионтного таксона (своего рода генералиста), способного одновременно эксплуатировать ресурсы нескольких лакун, пониженное конкурентное давление в них позволяет проникнуть в замкнутое до того сообщество. Особенно неожиданным оказывается то, что на четвертой фазе филоценогенеза сообщество приоткрывается для малоспециализированного эврибионтного иммигранта из экотона, оставаясь замкнутым для специалистов. Таким образом, мы справились со вторым экологическим парадоксом.
Необходимо подчеркнуть, что ресурсы экологических лакун и без такого «нувориши» не пропадают. Они полностью потребляются членами сообщества, но на них на данной фазе его эволюции не находится специализированного потребителя, а это, собственно, и означает, что конкурентное давление в лакунах понижено.
Появление экологических лакун — первый симптом вторичного понижения целостности, плотности упаковки адаптивных зон и экологических ниш и роста разнообразия малоспециализированных таксонов в сообществе. Возникновение лакун, как мы убедились, есть естественный результат противоречивых процессов прогрессивной специализации, которые, с одной стороны, постоянно наращивают конкурентоспособность ценофилов, а с другой — суживают фундаментальную нишу или адаптивную зону. Первое ведет к постоянному вытеснению менее специализированных форм, а второе не позволяет более специализированным вытеснителям полностью скомпенсировать конкурентное давление, которое осуществляли вытесненные формы по всей области бывших ресурсов. Одно без другого оказывается невозможным в силу принципа адаптивного компромисса.
Издержки специализации каждого таксона, в совокупности составляющие груз его инадаптивности, препятствует смене направления специализации (Раутиан, 1988). Структура адаптивных зон и экологических ниш, являясь первоначально продуктом экологической дифференцировки, получает посредством процессов филогенетической специализации отображение в организации членов соответствующих таксонов, что, в свою очередь, поддерживает и стимулирует дальнейшую специализацию в ранее избранном направлении даже на стадии вторичного понижения целостности сообщества.
5-я фаза филоценогенеза (см. рис. 4, Т6). Дальнейший филогенез специализированных ценофилов первой формации быстрее и медленнее, но с обязательностью закона будет вести: 1) к постепенному затуханию внутризональной конкуренции и вытеснению менее специализированных форм; тем самым 2) к понижению таксономического разнообразия ценофилов первой формации; 3) к постепенному расширению экологических лакун и образованию новых; 4) к проникновению в сообщество малоспециализированных форм из эко-тонов, которые по мере расширения имеющихся лакун и появления новых; 5) начинают новый цикл взаимозависимых процессов прогрессивной специализации, образуя группу ценофилов второй формации. Первым признаком последнего будет рост их таксономического разнообразия.
Проходя первые стадии специализации в зонах пониженного конкурентного давления, ценофилы второй формации на этой фазе филоценогенеза не вступают в активную конкуренцию с высоко-специализированными ценофилами первой формации. Такое на первый взгляд парадоксальное положение, как мы убедились, является следствием низкой толерантности высокоспециализированных форм по сравнению с формами, лишь начинающими свою прогрессивную специализацию.
6-я фаза филоценогенеза (см. рис. 4, Т7). Очаги специализации ценофилов второй формации — экологические лакуны — находятся на периферии адаптивные зон и экологических ниш ценофилов первой формации. Поэтому ценотические структуры ценофилов второй формации обычно не повторяют таковые ценофилов первой формации. Например, в Северной Америке адаптивная зона специализированных насекомоядов, представленная в неоген-современности тремя семействами, отсутствовала в палеогене, а ее ресурсы были распределены между древними группами насекомоядных, являющимися экологически хищниками — генералистами мелкого размерного класса. Напротив, адаптивные зоны хищников разных размерных классов в палеогене были обозначены значительно резче, чем в неоген-современности (см. рис. 5, а).
Это ведет ко все большей реорганизации сообщества и неизбежно понижает уровень экологической специализации ценофилов первой формации, вся филогенетическая специализация которых ориентирована на прежнюю структуру сообщества. Высокий груз инадаптивности и его оборотная сторона — низкая экологическая и особенно, филогенетическая толерантность — препятствуют смене направлений специализации и тем самым установлению нового соответствия экологической и филогенетической специализации. Это в конечном счете делает возможной конкуренцию с ними даже филогенетически менее специализированных ценофилов второй формации, что указывает на продолжающуюся реорганизацию сообщества и смену его эдификаторов.
Таким образом, мы получили решение третьего парадокса: как в недрах сформированного сообщества складываются условия формирования специалистов второй формации и реорганизации сообщества в целом.
7-я фаза филоценогенеза (см. рис. 4, T8). Реорганизация сообщества не обязательно завершается вымиранием всех ценофилов первой формации. Как правило, часть из них, обычно небольшая, обладающая сравнительно небольшим грузом инадаптивности, входит в новую преобразованную структуру сообщества, в которой продолжает свою прогрессивную специализацию, начатую на предшествующем этапе филоценогенеза (1—3 фазы). Ярким примером такого рода являются современные лошадиные (Equidae) — небольшая группа высокоспсциализированных непарнопалых, столь характерных и многочисленных в фаунах палеогена, нашедшая место в современной фауне.
Нормальная смена биоты. Описанный процесс реорганизации сообщества описывает процедуру нормальной постепенной смены биоты и преобразования структуры сообщества, не сопровождающихся значительным экологическим кризисом. Наблюдается лишь небольшой кризис устойчивости сообщества, сопровождающийся небольшим понижением разнообразия, понижением плотности упаковки адаптивных зон и понижением его замкнутости для иммигрантов. Небольшой масштаб нарушений проявляется в том, что иммигранты обладают незначительным уровнем специализации и происходят из экотонов, а не другого сформированного сообщества. Последнее имеет место лишь в случае коллизии биот, когда в сообщество проникают высокоспециализированные формы, вытесняющие автохтонные и ввергающие его в состояние экологического кризиса.
Низкая специализация и высокая толерантность иммигрантов из экотонов не приводит к быстрому радикальному преобразованию сообщества, структуру которого в процессе вселения продолжают определять прежние эдификаторы. Именно это обеспечивает затем постепенную и относительно безболезненную их интеграцию постепенно реорганизующимся сообществом.
Биота экотонов в момент назревания кризиса устойчивости сообщества, вызванного далеко зашедшими процессами прогрессивной специализации ценофилов, имеет ключевую роль. Проникновение ценофобов в сообщество со временем ведет к реставрации его устойчивости, плотности упаковки адаптивных зон и экологических ниш и к замкнутости для иммигрантов. Таким образом, существование экотонов и свойственных им ценофобов позволяет неоднократно проводить смену биоты и реорганизацию структуры сообщества, не ввергая его в состояние глубокого кризиса всякий раз, как издержки односторонней специализации его членов начинают подрывать его устойчивость. Это свидетельствует о глубокой исторической связи сообщества и окружающих его экотонов, своего рода равновесии между ними, существующем в каждый данный момент.
Историческая подготовка крупного кризиса. Длительный процесс относительно безконфликтного филоценогенеза, возможный благодаря периодической реставрации сообщества выходцами из экотонов, ведет к захвату все больших площадей этими процессами. Это связано: 1) с постоянной средообразующей работой сообщества, делающей доступной для жизни ценофилов все большие площади, и 2) с медленным, но постоянным наступлением сообщества на экотоны вследствие его большей устойчивости и преимуществ стратегии адаптации ценофилов по сравнению с ценофобами. К этому надо добавить периодическое обеднение экотонов в связи с переходом части ценофобов в сообщество в процессе его реставрации. Этот процесс нельзя назвать наступлением экотонов на сообщество, что имеет место только в условиях кризиса (простейшая модель такого кризиса—диаспорический субклимакс). В случае реставрации сообщество как бы «втягивает» в себя отдельные формы из экотонов. Наконец, низкая определенность условий жизни, прежде всего биотической среды, в экотонах и связанные с этим трудности формирования специфических адаптации ведут к более бедному таксономическому разнообразию биоты ценофобов и меньшей «сумме жизни» в экотонах.
Результатом этого, очевидно, является, хотя и не часто возникающая, неспособность ценофобов закрыть возникающие экологические лакуны и отреставрировагь сообщество. Тогда возникают условия для развития крупного экологического кризиса, подобного юрскому у наземных тетрапод, вызванному разрушением структуры прежнего сообщества.
Ввиду высокой толерантности и большей свободы эволюционного эксперимента в условиях отсутствия ценотических ограничений ценофобы, хотя и не часто, могут создавать очень ценные адаптации. Например, энтомофилию цветковых растений (Красилов, 1989). В условиях понижения устойчивости сообщества и роста экологических лакун такие ценофобы могут спровоцировать экологический кризис, образовав авангард наступления экотонов на сообщество, естественно на его слабые места — например, на ранние стадии сукцессии. Как подчеркивал В. В. Жерихин (1978—1987), перехват ресурсов пионерных стадий должен превратить возобновление поздних стадий сукцессии и тем самым вызвать гибель всей сукцессионной системы. Таков, очевидно, был сценарий биоценотического кризиса в середине мела.
Еще раз о причинах юрского кризиса. Нами указана лишь самая общая вероятная причина наступления крупного экологического кризиса эндогенной природы, в том числе и юрского. Однако мы далеки от мысли о полноте такого объяснения. Выяснение конкретных обстоятельств наступления юрского кризиса, подобных указанным для срединномелового, требуют дополнительных специальных исследований и потому откладываются до будущих публикаций.
ЛИТЕРАТУРА
Борисяк А. А. Курс исторической геологии. Л. и др.: Горгеонефтеиздат, 1934. 424 с.
Baxpамеев В. А. Юрские и меловые флоры и климаты Земли. М.: Наука, 1988. 212 с.
Вахрамеев В. А., Добрускина И. А., Заклинская Е. Д., Мечен С. В. Палеозойские и мезозойские флоры Евразии и фитогеография этого времени. М.: Наука, 1970. 424 с. (Тр. ГИН АН СССР; Вып. 208).
Вахрушев А. А. О закономерностях формирования сообществ на примере формирования комплекса птиц городов//Макроэволюция: (Материалы I Всесоюз. конф. по пробл. эволюции). М.: Наука, 1984. С. 148—149.
Вахрушев А. А. Синэкологические аспекты синантропизации//Изучение птиц СССР, их охрана и рациональное использование: Тез. докл. I съезда Всесоюз. орнитол. о-ва и IX Всесоюз. орнитол. конф. Л., 1986. Ч. 1. С. 117—118.
Вахрушев А. А. Особенности экологии птиц-синантропов в условиях большого города: Автореф. дис.... канд. биол. наук. М., 1988. 16 с.
Вахрушев А. А. Начальные этапы формирования сообществ на примере синантропизации птиц//Эволюционные исследования. Владивосток: ДВНЦ АН СССР, 1988. С. 34—36.
Вегенер А. Л. Происхождение континентов и океанов. Л.: Наука, 1984. 211 с.
Вернадский В. И. Биосфера//Соч. М.: Изд-во АН СССР, 1960. Т. 5. С. 7—102.
Длусский Г. М. Муравьи пустынь. М.: Наука, 1981. 230 с.
Дмитриев В. 10., Жерихин В. В. Изменения разнообразия семейств насекомых по данным методам накопления появлений//Мелоиой биоценотический кризис и эволюция насекомых. М.: Наука, 1988. С. 208—215.
Ефремов И. А. Тафономия и палеонтологическая летопись. М.: Изд-во АН СССР, 1950. 177 с. (Тр. НИН АН СССР; Т. 24, вып. 1).
Жерихин В. В. Развитие и смена меловых и кайнозойских фаунистических комплексов (трахейные и хелицеровые). М.: Наука, 1978. 198 с. (Тр. ПИИ АН СССР; Т 165).
Жерихин В. В. Использование палеонтологических данных в экологическом прогнозировании//Экологическое прогнозирование. М.:,Наука, 1979. С. 113—132.
Жерихин В. В. Класс Insecta: Насекомые//Развитие и смена беспозвоночных на рубеже мезозоя и кайнозоя: Мшанки, членистоногие, иглокожие. М.: Наука. 1980. С. 40—97.
Жерихин В. В. Биоценотическая регуляция эволюции//Палеонтол. жури. 1987. № 1 С. 3—12.
Жерихин В. В., Калугина И. С. Насекомые в экосистемах геологического прошлого//Историческое развитие класса насекомых. М.: Наука, 1980. С. 188—240 (Тр. НИН АН СССР; Т. 175).
Жерихин В. В., Калугина II. С. Ландшафты v сообщества//Юрскис континентальные биоценозы Южной Сибири и сопредельных территорий. М.: Наука, 1985. С. 137—183. (Тр. НИН АН СССР; Т. 213).
Жерихин В. В., Расницын А. П. Биоценотическая регуляция эволюционных процессов//Микро- и макроэволюция. Тарту, 1980. С. 77—81. Жерихин В. В., Сукачева И. Д. О меловых насекомоносных «янтарях» (ретинитах) севера Сибири//Вопросы палеонтологии насекомых: Докл. на XXIV Ежегод. чтении памяти II. А. Холодковского, 1971. Л.: Наука, 1973. С. 3—48.
Ивахненко М. Ф. Хвостатые амфибии из триаса и юры Средней Азии//Палеонтол.журн. 1978. № 3. С. 84—89.
Ивахненко М. Ф. Пермские парарептилии СССР. М.: Наука, 1987. 160 с. (Тр. ПИП» АН СССР; Т. 223).
Каландадзе Н. Н. О межконтинентальных связях фаун тетрапод в триасовом периоде//Палеонтол. журн. 1974. № 3. С. 75—86.
Каландадзе Н.Н., Раутиан А. С. О связях наземных тетрапод в каменноугольном, пермском и триасовом периодах: (Предварительные результаты анализа, проблемы и перспективы)//Жизнь на древних континентах, ее становление и развитие: Тез. докл. XXIII сес. Всесоюз. палеонтол. о-ва. Л.: ВСЕГЕИ, 1977. С. 33—34.
Каландадзе Н. И., Раутиан А. С. Специализация конкурирующих таксонов в связи со степенью стабильности экосистемы//Палеонтология и эволюция биосферы: Тез. докл. XXV сес. Всесоюз. палеонтол. о-ва. Л.: ВСЕГЕИ, 1979. С. 21—23.
Каландадзе Н. II., Раутиан А. С. К исторической зоогеографии наземных тетрапод конца палеозоя — начала мезозоя//ХХУ1 Междунар. геол. конгр.: Докл. сов. геологов: Палеонтология. Стратиграфия. М.: Наука, 1980. С. 93—102.
Каландадзе Н. Н., Раутиан А. С. Межконтинентальные связи наземных тетрапод и решение проблемы шотландской фауны Элгин//Жизнь на древних континентах, ее становление и развитие: Тр. XXIII сес. Всесоюз. палеонтол. о-ва. Л.: Наука, 1981. С. 123—133.
Каландадзе Н. Н., Раутиан А. С. Историческая зоогеография млекопитающих//Млекопитающие СССР: III съезд Всесоюз. териол. о-ва: Тез. докл. М.: Наука, 1982. Ч. 1. С. 38.
Каландадзе Н. Н., Раутиан А. С. Место Центральной Азии в зоогеографической истории мезозоя//Ископаемые рептилии Монголии. М.: Наука, 1983. С. 6—44. (Тр. ССМПЭ; Вып. 24).
Каландадзе Н.Н., Раутиан А. С. Специализация таксонов и устойчивость экосистемы//Макроэволюция: (Материалы I Всесоюз. конф. по пробл. эволюции). М.: Наука, 1984. С. 152—153.
Каландадзе Н. Н., Раутиан А. С. Историческая зоогеография млекопитающих//Оперативно-информационные материалы к I Всесоюз. совещ. по палеотериологии. М.: Паука, 1989. С. 28—30.
Каландадзе Н. Н., Раутиан А. С. Историческая зоогеография позднего триаса и реконструкция фауны Северной Африки//Палеонтол. журн. 1991. № 1. С. 3—14.
Калугина Н. С. Изменения подсемейственного состава хирономид (Diplera, Chironomidae) как показатель возможного эвтрофирования водоемов в конце мезозоя //Бюл. МОИП. Отд. биол. 1974. Т. 79, вып. 6. С. 45—56.
Калугина Н. С. Основные этапы экогенеза водных насекомых//Жизнь на древних I континентах, ее становление и развитие: Тез. докл. XXIII сес. Всесоюз. палеонтол. о-ва. Л.: ВСЕГЕИ, 1977. С. 36—37.
Катастрофы в истории Земли: Новый униформизм. М.: Мир, 1986. 471 с.
К расилов В. А. Филогения и систематика//Проблемы филогении и систематики: Материалы симпоз. Владивосток: ДВНЦ АН СССР» 1969. С. 12—30.
Красилов В. А. Происхождение и ранняя эволюция цветковых растений. М.: Наука 1989. 263 с.
Лебедев О. А. Первые четвероногие: поиски и находки//Природа. 1985 № 11 С. 26—36.
Мейен С. В. филогения высших растений и флорогенез//ХХУП Междунар. геол. конгр. М., 1984. Т. 2: Палеонтология. С. 75—80.
М ейен. С. В. Методы палеофлористических исследований//Современнан палеонтология. М.: Недра, 1988. Т. 2. С. 31—44.
Назаров В. И. Реконструкция ландшафтов Белоруссии по палеоэнтомологическим данным: (Антропоген). М.: Наука, 1984. 96 с. (Тр. НИН АН СССР; Т. 205).
Несов Л. А., Казнышкина Л. Ф., Черепанов Г. О. Динозавры — цератопсии и крокодилы мезозоя Средней Азии//Теоретические и прикладные аспекты современной палеонтологии: Тр. XXXIII сес. Всесоюз. палеонтол. о-ва. Л.: Наука, 1989. С. 144—154.
Одум Ю. Основы экологии. М.: Мир, 1975. 740 с.
Пономаренко А. Г. Эволюция экосистем, основные события//ХХУП Междунар. геол. конгр. М.: Наука, 1984. Т. 2: Палеонтология. С. 71—74.
Пономаренко А. Г. Насекомые в раннемеловых экосистемах Западной Монголии//Насекомые в раннемеловых экосистемах Западной Монголии. М.: Наука, 1986. С. 183—201. (Тр. ССМПЭ; Вып. 28).
Пономаренко А. Г., Калугина H. С. Общая характеристика насекомых местонахождения Манлай//Раннемеловое озеро Манлай. М.: Наука, 1980. С. 68—81. (Тр. ССМПЭ; Вып. 13).
Разумовский С. М. Закономерности динамики биоценозов. М.: Наука, 1981. 231 с
Расницын А. П. Живое существо как адаптивный компромисс//Макроэволюция: (Материалы I Всесоюз. конф. по пробл. эволюции). М.: Наука, 1984. С. 233—234.
Расницын А. П. Инадаптация и энадаптация/Палеонтол. журн. 1986. № 1. С. 3—7.
Расницын А. П. Темпы эволюции и эволюционная теория: (Гипотеза адаптивного компромисса) //Эволюция и биоценотические кризисы. М.: Наука, 1987. С. 46—64.
Расницын А. П. Проблема глобального кризиса наземных биоценозов в середине мелового периода//Меловой биоценотичсский кризис и эволюция насекомых. М.: Наука, 1988. С. 191—207.
Расницын А. П. Динамика семейств насекомых и проблема мелового биоценотического кризиса//Осадочная оболочка Земли в пространстве и времени: Стратиграфия и палеонтология. М.: Наука, 1989. С. 35—40.
Расницын А. П. История палеоэптомологии и история насекомых//Природа. 1990. № 6. С. 66—80.
Раутиан А. С. Уникальное перо из отложений юрского озера в хребте Каратау//Палеонтол. журн. 1978. № 4. С. 106—114.
Раутиан А. С. О конкурентных отношениях насекомоядных (Insectivora) и опоссумов (Didelphidae) в Северной Америке на протяжении кайнозоя//Палеонтология: Материалы науч. заседаний секции палеонтологии МОИП за 1977—1978 годы. М.: Наука, 1980. С. 98—99.
Раутиан А. С. Базальная радиация млекопитающих в свете данных исторической зоогеографии//Бюл. МОИП. Отд. геол. 1982. Т. 57, вып. 3. С. 122.
Раутиан А. С. Палеонтология как источник сведений о закономерностях и факторах эволюции//Современная палеонтология. М.: Недра, 1988. Т. 2. С. 76—118.
Раутиан А. С., Каландадзе H. H. Некоторые эволюционные уроки исторической зоогеографии//Макроэволюция: (Материалы I Всесоюз. конф. по пробл. эволюции). М.: Наука, 1984. С. 166—167.
Раутиан А. С., Каландадзе H. H. Дефицит крупных хищников в древних фаунах млекопитающих//0перативно-информационные материалы к I Всесоюз. совещ. по палеотериологии. М.: Наука, 1989. С. 47—49.
Рид Г., Уотсон Дж. История Земли: Поздние стадии истории Земли. Л. Недра, 1981. 408 с.
Родендорф Б. Б., Жерихин В. В. Палеонтология и охрана природы//Природа. 1974. № 5. С. 82—91.
Руженцев В. Е. Принципы систематики, система и филогения палеозойских аммоноидей. М.: Изд-во АН СССР, 1960. 331 с. (Тр. ПИИ АН СССР; Т. 83).
Северцов А. С. Факторы, ограничивающие адаптивную зону филогенеза амфибий//Журн. общ. биологии. 1978. Т. 39, № 1. С. 66—75.
Симпсон Дж. Г. Темпы и формы эволюции. М.: Изд-во иностр. лит., 1948. 358 с.
Соколов Б. С. Проблемы эволюции биосферы: (Историзм и актуализм в проблемах глобальной экологии)//Вести. АН СССР. 1988. № 11. С. 17—21.
Страхов H. М. Основы исторической геологии. М.; Л.: Госгеолтехиздат, 1948. Т. 1. 253s с.; Т. 2. 396 с.
Татаринов Л. П. Морфологическая эволюция териодонтов и общие вопросы филогенетики. М.: Наука, 1976. 257 с.
Татаринов Л. П. "Очерки по теории эволюции. М.: Наука, 1987. 251 с.
Хэллем А. Юрский период. Л.: Недра, 1978. 272 с.
Шишкин М. А. Лабиринтодонт из юры МНР//Палеонтол. журн. 1990. № 4.
Шмальгаузен И. И. Факторы эволюции: Теория стабилизирующего отбора. М Наука, 1968. 451 с.
Шмальгаузен И. И. Избранные труды. Организм как целое в индивидуальном и историческом развитии. М.: Наука, 1982. 383 с.
Шмальгаузен И. И. Пути и закономерности эволюционного процесса. М.: Наука, 1983. 360.с.
Элтон Ч. Экология нашествий животных и растений. М.: Изд-во иностр. лит., 1960.
Эшби У. Р. Введение в кибернетику. М.: Изд-во иностр. лит., 1959, 432 с.
Эшби У. Р. Конструкция мозга: Происхождение адаптивного поведения. М.: Изд-во иностр. лит., 1962. 398 с.
Benton М. J. Mass extinclion among families of non-marine tetrapods//Nature 1985 Vol 306, N 6031. Р. 811—814.
Benton М.J. Mass extinction among families of non-marine tctrapods: The data//Mem Soc. geol. France. 1987. N 150. P. 21—32.
Benton М.J. Mass extinction on the fossil record of reptiles paraphyly, patchiness and periodicity//Extinction and survival in the fossil record. Oxford: Clarendon press 1988. (Syst. Assoc. Spec.; N 34). P. 269—294.
Benton М. ]. Patterns of evolution and exitinclion in vertebrates//Evolution and the fossil record. L.: Belhaven press, 1989. P. 218—241.
Carroll R. L. Vertebrate paleontology and evolution. N. Y.: Freeman, 1988. 698 p.
Сarroll R. L., Caskill P. The order Microsauria. Philadelphia: Amer. Philos. Soc., 1978 211 p.
Соре Е. D. The primary factors of organic evolution. Chicago, 1904.
Milner A. Late extinctions of Amphibians//Nature. 1989. Vol. 338. P. 117.
Muller A. H. Lehrbuch der Palaozoologie. Jena: Fischer, 1976. Bd. 1.
Nessov L. A. Late mezozoic amphibians and lizards of Soviet Middle Asia//Acta zool cracov. 1988. Vol. 34, N 14. P. 475—486.
Paul G. S. Predatory Dinosauns of the world. N. Y. etc.: Simon and Schuster, 1988. 464 p.
Schindewolf 0. Grundlagen der Palaontologie: Geologische Zeitmessung. Organische Stammesentwicklung. Biologische Systematik. Stuttgart: Springer, 1950.
Wallace A. R. Die geographische Verbreitung der Thiere. Dresden: Zahn, 1876. Bd. 1 579 S.; Bd. 2. 658 S.
|
|
|
|
|
Дата добавления: 2015-05-29; Просмотров: 617; Нарушение авторских прав?; Мы поможем в написании вашей работы!