КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Лекция 32. Сенсорные системы. Физиология анализаторов
|
|
|
|
32. 1. Рецептор: понятие, функция, классификация рецепторов, свойства и их особенности, механизм возбуждения рецепторов.
Сенсорная система выполняет следующие основные функции, или операции, с сигналами: 1) обнаружение; 2) различение; 3) передачу и преобразование; 4) кодирование; 5) детектирование признаков; 6) опознание образов.
Обнаружение и первичное различение сигналов обеспечивается рецепторами, а детектирование и опознание сигналов — нейронами коры больших полушарий. Передачу, преобразование и кодирование сигналов осуществляют нейроны всех слоев сенсорных систем.
При всем разнообразии стимулов и сенсорных систем все системы имеют одинаковый план строения. Каждая сенсорная система состоит из периферической части – рецепторов, проводниковой – нервных путей и подкорковых нервных центров, корковой части – в ней происходит окончательный анализ информации, поступившей от периферических рецепторов и нервных центров коры больших полушарий.
Классификация рецепторов. В практическом отношении наиболее важное значение имеет психофизиологическая классификация рецепторов по характеру ощущений, возникающих при их раздражении. Согласно этой классификации, у человека различают зрительные, слуховые, обонятельные, вкусовые, осязательные рецепторы, термо-, проприо- и вестибулорецепторы (рецепторы положения тела и его частей в пространстве) и рецепторы боли.
Существуют рецепторы внешние (экстерорецепторы) и внутренние (интерорецепторы). К экстерорецепторам относятся слуховые, зрительные, обонятельные, вкусовые, осязательные. К интерорецепторам относятся вестибуло- и проприорецепторы (рецепторы опорно-двигательного аппарата), а также висцерорецепторы (сигнализирующие о состоянии внутренних органов).
|
|
|
По характеру контакта со средой рецепторы делятся на дистантные, получающие информацию на расстоянии от источника раздражения (зрительные, слуховые и обонятельные), и контактные — возбуждающиеся при непосредственном соприкосновении с раздражителем (вкусовые, тактильные).
В зависимости от природы раздражителя, на который они оптимально настроены, рецепторы могут быть разделены на фоторецепторы, механорецепторы, к которым относятся слуховые, вестибулярные рецепторы, и тактильные рецепторы кожи, рецепторы опорно-двигательного аппарата, барорецепторы сердечно-сосудистой системы; хеморецепторы, включающие рецепторы вкуса и обоняния, сосудистые и тканевые рецепторы; терморецепторы (кожи и внутренних органов, а также центральные термочувствительные нейроны); болевые (ноцицептивные) рецепторы.
Все рецепторы делятся на первично-чувствующие и вторично-чувствующие. К первым относятся рецепторы обоняния, тактильные и проприорецепторы. Они различаются тем, что преобразование энергии раздражения в энергию нервного импульса происходит у них в первом нейроне сенсорной системы. К вторично-чувствующим относятся рецепторы вкуса, зрения, слуха, вестибулярного аппарата. У них между раздражителем и первым нейроном находится специализированная рецепторная клетка, не генерирующая импульсы. Таким образом, первый нейрон возбуждается не непосредственно, а через рецепторную (не нервную) клетку.
Общие механизмы возбуждения рецепторов. Рецепторы представляют собой клетки, различающие естественные раздражители и посылающие информацию о них в ЦНС. Стимуляция рецептора вызывает в дендритах изменение потенциала покоя в сторону деполяризации. При действии стимула на рецепторную клетку происходит преобразование энергии внешнего раздражения в рецепторный сигнал, или трансдукция сенсорного сигнала. Этот процесс включает в себя три основных этапа:
|
|
|
1) взаимодействие стимула, т. е. молекулы пахучего или вкусового вещества (обоняние, вкус), кванта света (зрение) или механической силы (слух, осязание) с рецепторной белковой молекулой, которая находится в составе клеточной мембраны рецепторной клетки;
2) внутриклеточные процессы усиления и передачи сенсорного стимула в пределах рецепторной клетки; и
3) открывание находящихся в мембране рецептора ионных каналов, через которые начинает течь ионный ток, что, как правило, приводит к деполяризации клеточной мембраны рецепторной клетки (возникновению так называемого рецепторного потенциала).
В первично-чувствующих рецепторах этот потенциал действует на наиболее чувствительные участки мембраны, способные генерировать потенциалы действия — электрические нервные импульсы. Во вторично-чувствующих рецепторах рецепторный потенциал вызывает выделение квантов медиатора из пресинаптического окончания рецепторной клетки. Медиатор (например, ацетилхолин), воздействуя на постсинаптическую мембрану первого нейрона, изменяет ее поляризацию (генерируется постсинаптический потенциал). Постсинаптический потенциал первого нейрона сенсорной системы называют генераторным потенциалом, так как он вызывает генерацию импульсного ответа.
В первично-чувствующих рецепторах рецепторный и генераторный потенциалы — одно и то же.
Генерация возбуждения в рецепторах. Возникновение рецепторного потенциала обусловлено повышением Na+-проводимости дендритов. Возникающее в них возбуждение электротонически распространяется к соме – происходит преобразование или первичная трансформация стимула в рецепторный потенциал. Поэтому рецептор представляет собой преобразователь, датчик. Возбуждение в форме рецепторного потенциала охватывает только сому. В аксоне же первичных рецепторов, начиная от аксонного холмика – места отхода аксона от сомы – происходит трансформация этого возбуждения в серию потенциалов действия.
Очень важно, что после первого потенциала действия мембрана аксона гиперполяризуется существенно ниже уровня потенциала покоя. Благодаря этому обстоятельству Na+-каналы после инактивации восстанавливаются настолько, что фаза деполяризации, наступающая после первого следового гиперполяризационного потенциала, вновь достигает порогового значения, достаточного для генерации следующего потенциала действия. Следовательно, гиперполяризационный следовой потенциал служит основой формирования ритмического возбуждения нервного волокна.
|
|
|
Во вторичных рецепторах возникает только рецепторный потенциал, а серия потенциалов действия формируется в терминалях афферентной нервной клетки, образующей контакт с рецептором. В частности, зрительные и слуховые рецепторы являются вторичными.
Когда генераторный потенциал (ГП) достигает критической величины, он вызывает разряд афферентных импульсов в ближайшем перехвате Ранвье. Частота разряда прямо пропорциональна величине ГП (логарифмическая зависимость, соответствующая закону Вебера-Фехнера). Ощущение тоже увеличивается пропорционально логарифму силы раздражения. Новокаин прерывает поток этих импульсов, с чем связан его аналгетический эффект.
Адаптация рецепторов - общее свойство всех рецепторов, заключающееся в приспособлении к силе раздражителя. Она проявляется в снижении чувствительности к постоянно действующему раздражителю. Человек "привыкает" к действию постоянных раздражителей - запаху, давлению одежды, звуку часов и т.п. и перестает замечать их. При адаптации снижается величина генераторного потенциала и частота импульсов, проходящих по афферентному нерву.
Есть медленно адаптирующиеся рецепторы (болевые) и быстро адаптирующиеся (глаз). Не адаптируются (или почти не адаптируются) только вестибуло- и проприорецепторы. Когда действие постоянного раздражителя прекращается, адаптация исчезает и чувствительность рецептора повышается (эффект возбуждения после торможения).
Когда начинается действие какого-либо стимула рецептор реагирует на него очень энергично. По мере продолжения стимуляции рецептор адаптируется к нему, и активность в сенсорном волокне снижается до более низкого уровня. При коротких и периодических предъявлениях стимула рецептор каждый раз реагирует на него полностью, без адаптации.
|
|
|
Медленно адаптирующиеся рецепторы служат для контроля за длительно сохраняющимися стимулами, например, степенью растяжения мышц, концентрацией Н+. Быстро адаптирующиеся рецепторы свойственны сенсорным системам, регистрация стимулов в которых происходит с высокой чувствительностью и высоким временным разрешением.
Различение сигналов. Важная характеристика сенсорной системы — способность замечать различия в свойствах одновременно или последовательно действующих раздражителей. Различение начинается в рецепторах, но в этом процессе участвуют нейроны всей сенсорной системы. Оно характеризует то минимальное различие между стимулами, которое сенсорная система может заметить (дифференциальный, или разностный, порог).
Зависимость силы ощущения от силы раздражения (закон Вебера—Фехнера) выражается формулой: E=a∙logI +b, где Е — величина ощущения, I — сила раздражения, а и b — константы, различные для разных модальностей стимулов. Согласно этой формуле, ощущение увеличивается пропорционально логарифму интенсивности раздражения.
Абсолютную чувствительность сенсорной системы измеряют порогом реакции. Чувствительность и порог — обратные понятия: чем выше порог, тем ниже чувствительность, и наоборот. Обычно принимают за пороговую такую силу стимула, вероятность восприятия которого равна 0,5 или 0,75 (правильный ответ о наличии стимула в половине или в 3/4 случаев его действия). Более низкие значения интенсивности считаются подпороговыми, а более высокие — надпороговыми. Оказалось, что и в подпороговом диапазоне реакция на сверхслабые раздражители возможна, но она неосознаваема (не доходит до порога ощущения). Так, если снизить интенсивность вспышки света настолько, что человек уже не может сказать, видел он ее или нет, от его руки можно зарегистрировать неощущаемую кожно-гальваническую реакцию на данный сигнал. Чувствительность рецепторных элементов к адекватным раздражителям, к восприятию которых они эволюционно приспособлены, предельно высока. Так, обонятельный рецептор может возбудиться при действии одиночной молекулы пахучего вещества, фоторецептор — одиночным квантом света. Чувствительность слуховых рецепторов также предельна: если бы она была выше, мы слышали бы постоянный шум из-за теплового движения молекул.
Выше упоминалось о различении силы раздражителей. Пространственное различение основано на распределении возбуждения в слое рецепторов и в нейронных слоях. Так, если два раздражителя возбудили два соседних рецептора, то различение этих раздражителей невозможно и они будут восприняты как единое целое. Необходимо, чтобы между двумя возбужденными рецепторами находился хотя бы один невозбужденный. Для временного различения двух раздражений необходимо, чтобы вызванные ими нервные процессы не сливались во времени и чтобы сигнал, вызванный вторым стимулом, не попадал в рефрактерный период от предыдущего раздражения.
Амплитуда (интенсивность) стимула кодируется в виде частоты импульсов или потенциалов действия, направляющихся от рецептора в ЦНС. Повышение амплитуды стимула при условии ее надпорогового значения соответственно повышает частоту потенциалов действия.
32.2. Анализаторы (И.П. Павлов): понятие, классификация анализаторов, три отдела анализаторов и их значение, принципы построения корковых отделов анализаторов.
Анализатором, по И. П. Павлову, называют часть нервной системы, состоящую из воспринимающих элементов — сенсорных рецепторов, получающих стимулы из внешней или внутренней среды, нервных путей, передающих информацию от рецепторов в мозг, и тех частей мозга, которые перерабатывают эту информацию.
Методы изучения сенсорных систем. Для изучения сенсорных систем используют электрофизиологические, нейрохимические, поведенческие и морфологические исследования на животных, психофизиологический анализ восприятия у здорового и больного человека, методы картирования его мозга. Сенсорные функции также моделируют и протезируют.
Моделирование сенсорных функций позволяет изучать на биофизических или компьютерных моделях такие функции и свойства сенсорных систем, которые пока недоступны для экспериментальных методов.
Основные принципы строения анализаторов. Основными общими принципами построения сенсорных систем высших позвоночных животных и человека являются следующие:
1) Многослойность, т. е. наличие нескольких слоев нервных клеток, первый из которых связан с рецепторами, а последний - с нейронами моторных областей коры большого мозга.
2) Многоканальность сенсорной системы, т. е. наличие в каждом слое множества (от десятков тысяч до миллионов) нервных клеток, связанных с множеством клеток следующего слоя. Наличие множества таких параллельных каналов обработки и передачи информации обеспечивает сенсорной системе точность и детальность анализа сигналов и большую надежность;
3) Наличие сенсорных воронок. Разное число элементов в соседних слоях формирует «сенсорные воронки». Так, в сетчатке глаза человека насчитывается 130 млн фоторецепторов, а в слое ганглиозных клеток сетчатки нейронов в 100 раз меньше («суживающаяся воронка»). На следующих уровнях зрительной системы формируется «расширяющаяся воронка»: число нейронов в первичной проекционной области зрительной области коры в тысячи раз больше, чем ганглиозных клеток сетчатки. В слуховой и в ряде других сенсорных систем от рецепторов к коре большого мозга идет «расширяющаяся воронка». Физиологический смысл «суживающейся воронки» заключается в уменьшении избыточности информации, а«расширяющейся» — в обеспечении дробного и сложного анализа разных признаков сигнала; дифференциация сенсорной системы по вертикали и по горизонтали.
4) Вертикальная и горизонтальная дифференциация. Дифференциация по вертикали заключается в образовании отделов, каждый из которых состоит из нескольких нейронных слоев. Таким образом, отдел представляет собой более крупное морфофункциональное образование, чем слой нейронов. Каждый отдел (например, обонятельные луковицы, кохлеарные ядра слуховой системы или коленчатые тела) осуществляет определенную функцию. Дифференциация по горизонтали заключается в различных свойствах рецепторов, нейронов и связей между ними в пределах каждого из слоев. Так, в зрении работают два параллельных нейронных канала, идущих от фоторецепторов к коре большого мозга и по-разному перерабатывающих информацию, поступающую от центра и от периферии сетчатки глаза.
Общие принципы формирования анализаторов. Общим для большинства проводящих путей анализаторов является то, что они перед попаданием в ядерные зоны коры отдают коллатерали ретикулярной формации и взаимодействуют с ней, а также проходят через таламус.
Корковым представительством анализаторов являются первичные и вторичные поля, преимущественно расположенные в затылочных, постцентральных и височных отделах второго блока (блока приема, переработки и хранения экстероцептивной информации) мозга.
Все анализаторные системы функционируют на основе следующих общих принципов:
1) анализа информации с помощью специальных нейронов-детекторов;
2) параллельной многоканальной переработки информации, обеспечивающей ее надежность;
3) селекции информации в промежутке от рецептора до проекционного поля;
4) последовательного усложнения переработки информации от уровня к уровню;
5) целостной представленности сигнала в ЦНС во взаимосвязи с другими сигналами;
6) реализации принципов повышения надежности обработки разных признаков сигнала.
Основу корковых отделов анализаторов составляют первичные или проекционные зоны коры (поля), выполняющие узкоспециализированную функцию отражения только стимулов одной модальности. Их задача — идентифицировать стимул по его качеству и сигнальному значению, в отличие от периферического рецептора, который дифференцирует стимул лишь по его физическим или химическим характеристикам. Основная функция первичных полей — тончайшее отражение свойств внешней и внутренней среды на уровне ощущения.
Все первичные корковые поля характеризуются топическим (экранным) принципом организации, согласно которому любому участку рецепторной поверхности соответствует определенный участок в первичной коре (по принципу «точка в точку»), что и дало основание назвать первичную кору проекционной. Величина зоны представительства того или иного рецепторного участка в первичной норе зависит от функциональной значимости этого участка, а не от его фактического размера.
К числу первичных относятся поля: 17-е (для зрения). 3-е (для кожно-кинестетической чувствительности) и 41-е (для слуха). Экстероцепторная информация в эти участки мозга попадает после прохождения через релейные ядра таламуса.
Вторичные поля представляют клеточные структуры, морфологически и функционально как бы надстроенные над проекционными. В них происходит последовательное усложнение процесса переработки информации, чему способствует предварительное проведение афферентных импульсов через ассоциативные ядра таламуса. Вторичные поля обеспечивают превращение соматотропических импульсов в такую функциональную организацию, которая на уровне психики эквивалентна процессу восприятия.
На поверхности мозга вторичные поля граничат с проекционными или окружают их. Номера вторичных полей: 18,19 — для зрения, 1,2 и частично 5 — для кожно-кинестетической чувствительности, 42 и 22 — для слуха. Первичные и вторичные поля относятся к ядерным зонам анализаторов, расположенных на трех пространственных полюсах заднего мозга – затылочного, теменного и височного соответственно.
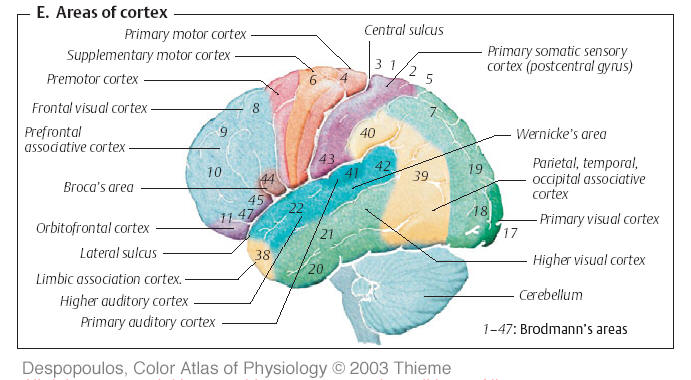
Рис.56. Поля коры головного мозга.
Третичные поля (ассоциативные, зона перекрытия) принимают на себя наиболее сложную функциональную нагрузку. Они находятся вне ядерных зон и в основном расположены в промежутке между вторичными полями или по их периметру. Большая и важнейшая часть третичных полей формируется на границе теменного, затылочного и височного отделов, оказываясь равноудаленной от каждого из указанных полюсов, и не имеет непосредственного выхода на периферию. Их функции почти полностью сводятся к интеграции возбуждений, приходящих от вторичной коры всего комплекса анализаторов. Работа третичных зон своим психологическим эквивалентом имеет восприятие мира во всей полноте и комбинации пространственных, временных и интенсивностных характеристик внешней среды. Все это дает основание рассматривать их как аппарат межанализаторных синтезов.
|
|
|
|
|
Дата добавления: 2015-05-26; Просмотров: 6075; Нарушение авторских прав?; Мы поможем в написании вашей работы!