КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Торможение в ЦНС
|
|
|
|
Торможение - это активный нервный процесс, результатом которого является прекращение или ослабление возбуждения. Торможение вторично относительно процесса возбуждения, так как всегда возникает как следствие возбуждения.
Торможение в ЦНС открыл И. М. Сеченов (1863). В опыте на таламической лягушке он определял латентное время сгибатель-ного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Было показано, что латентное время рефлекса значительно увеличивается, если на зрительный бугор предварительно положить кристаллик поваренной соли. Открытие И. М. Сеченова послужило толчком для дальнейших исследований торможения в ЦНС, при этом было открыто два механизма торможения: пост- и пресинаптическое.
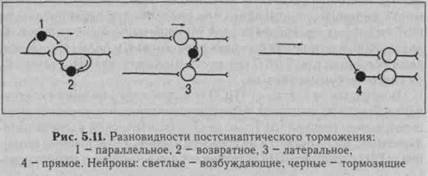
А. Постсинаптическое торможение возникает на постси-наптических мембранах нейрона в результате гиперполяризационного постсинаптическрго потенциала, уменьшающего возбудимость нейрона, угнетающего его способность реагировать на возбуждающие влияния. По этой причине вызванный гиперполяризационный потенциал был назван тормозным постсинаптиче-ским потенциалом, ТПСП '(см. рис. 5.6). АмплитудаТПСП 1-5 мВ, он способен суммироваться.
Возбудимость клетки от ТПСП (гиперполяризационного постси-наптического потенциала) уменьшается потому, что увеличивается пороговый потенциал (МО, так как Екр (критический уровень деполяризации, КУД) остается на прежнем уровне, а мембранный потенциал (Е) возрастает. ТПСП возникает под влиянием и аминокисло-
 ты глицина, и ГАМК - гамма-аминомасляной кислоты. В спинном мозге глицин выделяется особыми тормозными клетками (клетками Реншоу) в синапсах, образуемых этими клетками на мембране нейрона-мишени. Действуя на ионотропный рецептор постсинапти-ческой мембраны, глицин увеличивает ее проницаемость для СГ, при этом СГ поступает в клетку согласно концентрационному градиенту вопреки электрическому градиенту, в результате чего развивается гиперполяризация. В безхлорной среде тормозная роль глицина не реализуется. Ареактивность нейрона к возбуждающим импульсам является следствием алгебраической суммации ТПСП и ВПСП, в связи с чем в зоне аксонного холмика не происходит деполяризации мембраны до критического уровня. При действии ГАМК на постсинаптическую мембрану ТПСП развивается в результате входа СГ в клетку или выхода К+ из клетки. Концентрационные градиенты ионов К+ в процессе развития торможения нейронов поддерживаются Ыа/К-помпой, ионов СГ - СГ-помпой. Разновидности постсинаптического торможения представлены на рис. 5.11.
ты глицина, и ГАМК - гамма-аминомасляной кислоты. В спинном мозге глицин выделяется особыми тормозными клетками (клетками Реншоу) в синапсах, образуемых этими клетками на мембране нейрона-мишени. Действуя на ионотропный рецептор постсинапти-ческой мембраны, глицин увеличивает ее проницаемость для СГ, при этом СГ поступает в клетку согласно концентрационному градиенту вопреки электрическому градиенту, в результате чего развивается гиперполяризация. В безхлорной среде тормозная роль глицина не реализуется. Ареактивность нейрона к возбуждающим импульсам является следствием алгебраической суммации ТПСП и ВПСП, в связи с чем в зоне аксонного холмика не происходит деполяризации мембраны до критического уровня. При действии ГАМК на постсинаптическую мембрану ТПСП развивается в результате входа СГ в клетку или выхода К+ из клетки. Концентрационные градиенты ионов К+ в процессе развития торможения нейронов поддерживаются Ыа/К-помпой, ионов СГ - СГ-помпой. Разновидности постсинаптического торможения представлены на рис. 5.11.
|
|
|
|
Б. Пресинаптическое торможение развивается в преси-наптических окончаниях. При этом мембранный потенциал и возбудимость исследуемых нейронов не изменяются либо регистрируется низкоамплитудный ВПСП, недостаточный для возникновения ПД (рис. 5.12). Возбуждение блокируется в преси» наптических окончаниях вследствие деполяризации их. В очаге деполяризации нарушается процесс распространения возбуждения, следовательно, поступающие импульсы, не имея возможности пройти зону деполяризации в обычном количестве и обычной амплитуды, не обеспечивают выделение медиатора в синаптическую щель в достаточном количестве, поэтому нейрон не возбуждается, его функциональное состояние, естественно, остается неизменным. Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные клетки, аксоны которых образу-

ют синапсы на пресинаптических окончаниях аксона-мишени (см. рис 5.12). Торможение (деполяризация) после одного афферентного залпа продолжается 300-400 мс, медиатором является гамма-аминомасляная кислота (ГАМК), которая действует на ГАМК-рецепторы.
|
|
|
Деполяризация является следствием повышения проницаемости для СГ, в результате чего он выходит из клетки согласно электрическому градиенту. Это доказывает, что в составе мембран пресинаптических терминалей имеется хлорный насос, обеспечивающий транспорт СГ внутрь клетки вопреки электрическому градиенту.
Разновидности пресинаптического торможения изучены недостаточно. По-видимому, имеются те же варианты, что и для постсинаптического торможения. В частности, на рис. 5.12 представлено параллельное и латеральное пресинаптическое торможение. Однако возвратное пресинаптическое торможение на уровне спинного мозга (по типу возвратного постсинаптического торможения) у млекопитающих обнаружить не удалось, хотя у лягушек
оно выявлено.
В реальной действительности взаимоотношения возбуждающих и тормозных нейронов значительно сложнее, чем представлено на рис. 5.11 и 5.12, тем не менее все варианты пре- и постсинаптического торможений можно объединить в две группы: 1) когда блокируется собственный путь самим распространяющимся возбуждением с помощью вставочных тормозных клеток (параллельное и возвратное торможение) и 2) когда блокируются другие нервные элементы под влиянием импульсов от соседних возбуждающих нейронов с включением тормозных клеток (латеральное и прямое торможения). Поскольку тормозные клетки сами могут быть заторможены другими тормозными нейронами (торможение торможения), это может облегчить распространение возбуждения.
В. Роль торможения.
1. Оба известных вида торможения со всеми их разновидностями выполняют охранительную роль. Отсутствие торможения привело бы к истощению медиаторов в аксонах нейронов и прекращению деятельности ЦНС.
2. Торможение играет важную роль в обработке поступающей в ЦНС информации. Особенно ярко выражена эта роль у пре-синаптического торможения. Оно более точно регулирует процесс возбуждения, поскольку этим торможением могут быть заблокированы отдельные нервные волокна. К одному возбуждающему нейрону могут подходить сотни и тысячи импульсов по разным терми-налям. Вместе с тем, число дошедших до нейрона импульсов определяется пресинаптическим торможением. Торможение латеральных путей обеспечивает выделение существенных сигналов, из фона. Блокада торможения ведет к широкой иррадиации возбуждения и судорогам (например, при выключении пресинаптического торможения бикукулином).
|
|
|
3. Торможение является важным фактором обеспечения координационной деятельности ЦНС.
|
|
|
|
|
Дата добавления: 2015-05-26; Просмотров: 1581; Нарушение авторских прав?; Мы поможем в написании вашей работы!