КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Понятие о рефлексе и рефлекторной дуге
СТРОЕНИЕ НЕРВНОЙ ТКАНИ.
КЛЮ
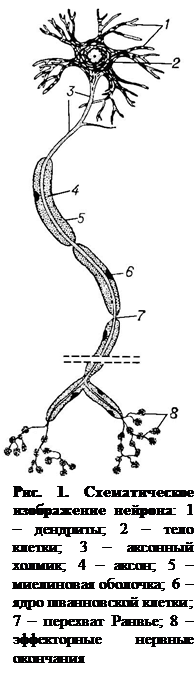 1. Строение нейрона. В состав нервной ткани входят высокоспециализированные нервные клетки, называемые нейронами, и клетки нейроглии. Последние тесно связаны с нервными клетками и выполняют опорную, секреторную и защитную функции.
1. Строение нейрона. В состав нервной ткани входят высокоспециализированные нервные клетки, называемые нейронами, и клетки нейроглии. Последние тесно связаны с нервными клетками и выполняют опорную, секреторную и защитную функции.
Нейрон – это основная структурная и функциональная единица нервной системы. Нейрон способен генерировать нервные импульсы, проводить возбуждение, принимать, обрабатывать, кодировать, хранить информацию, организовывать реакции на раздражения, устанавливать контакты с другими нейронами, клетками органов и выделять биологически активные вещества. В нем различают тело клетки, или сому, один мало ветвящийся отросток – аксон и много (от 1 до 1000) сильно ветвящихся отростков – дендритов (рис. 1).
Функционально в нейроне выделяют следующие части: воспринимающую – дендриты, мембрана сомы нейрона; интегративную – сома с аксонным холмиком; передающую – аксонный холмик с аксоном.
Тело нейрона в различных отделах нервной системы имеет разную величину (диаметр его колеблется от 4 до 130 мкм) и форму (округлую, уплощенную, многоугольную, овальную). Оно покрыто сложно устроенной мембраной и содержит органоиды, свойственные любой другой клетке: в цитоплазме находятся ядро с одним или несколькими ядрышками, митохондрии, рибосомы, аппарат Гольджи, эндоплазматическая сеть и др.
Характерной особенностью строения нервной клетки является наличие гранулярного ретикулума с большим количеством рибосом (базофильное вещество, вещество Ниссля, тигроидное вещество или тигроид) и нейрофибрилл. С рибосомами в нервных клетках связывают высокий уровень обмена веществ, синтез белка и РНК. Длительное возбуждение нейрона приводит к исчезновению в клетке базофильного вещества, а значит, и к прекращению синтеза специфического белка. У новорожденных нейроны лобной доли коры большого мозга не имеют базофильного вещества. В то же время в структурах, обеспечивающих жизненно важные рефлексы – спинном мозге, мозговом стволе, нейроны содержат большое количество базофильного вещества.
Нейрофибриллы представляют собой тончайшие волоконца, пересекающие тело клетки во всех направлениях и продолжающиеся в отростки. Они образуют в цитоплазме сложную трехмерную опорно-сократительную сеть, играющую важную роль в функционировании нейрона, хранении и передаче информации, а также транспорте веществ (в первую очередь медиаторов) внутри клетки и по ее отросткам.
Тело нейрона (сома), помимо информационной, выполняет трофическую функцию по отношению к своим отросткам и их синапсам. Перерезка аксона или дендрита ведет к гибели отростков, лежащих дистальнее перерезки, а, следовательно, и синапсов этих отростков. Сома обеспечивает также рост дендритов и аксона.
Аксон данной клетки имеет постоянный диаметр, в большинстве случаев одет в миелиновую оболочку, образованную из глии, от него отходят отдельные отростки – коллатерали. Аксон имеет разветвленные окончания. В окончаниях находятся митохондрии и секреторные образования. Длина аксона составляет от 0,5 мм до метра и более, его диаметр колеблется от сотых долей микрона (мкм) до 10 мкм. На протяжении первых 50 – 100 мкм от тела клетки аксон не покрыт миелиновой оболочкой. Прилегающий к нему участок тела клетки называют аксонным холмиком. В области аксонного холмика возбудимость мембраны нейрона наибольшая, именно здесь происходит генерация нервного импульса данного нейрона.
Аксон передает возбуждение от одного нейрона к другому или рабочему органу. Один аксон может контактировать с 5 тыс. нервных клеток и создавать до 10 тыс. контактов. Обработка поступившей в него информации происходит путем сложных нейрохимических перестроек белковых молекул в нейротрубочках тела нейрона, в результате в аксонном холмике генерируется возбуждение. Участок аксона, не покрытый миелиновой оболочкой, вместе с аксонным холмиком называют начальным сегментом аксона.
По дендритам возбуждение от рецепторов или других нейронов передается к телу клетки. Длина дендрита, как правило, не превышает 300 мкм (хотя длина дендрита афферентного нейрона может достигать метра и более), а диаметр его составляет – 5 мкм. На дендритах имеются боковые отростки (шипики), которые увеличивают их поверхность и являются местами наибольших контактов с другими нейронами. Чем сложнее функция нервной системы, чем больше разных анализаторов посылают информацию к данной структуре, тем больше «шипиков» на дендритах нейронов. Максимальное количество их содержится на пирамидных нейронах двигательной зоны коры большого мозга и достигает нескольких тысяч. За счет «шипиков» воспринимающая поверхность нейрона значительно возрастает и может достигать, например у клеток Пуркинье, 250 000 мкм. Если данный «шипик» или группа «шипиков» длительное время перестает получать информацию, то они исчезают.
Основной особенностью обмена веществ в нейроне является высокая скорость обмена и преобладание аэробных процессов. Потребность мозга в кислороде очень велика. Хотя вес мозга по отношению к весу тела составляет всего 2%, потребление кислорода мозгом достигает в состоянии покоя у взрослых людей 25% от общего его потребления организмом, а у маленьких детей – 50%. Энерготраты мозга составляют 1/6 – 1/8 суточных затрат организма человека.
Все нейроны ЦНС соединяются друг с другом в основном в одном направлении: разветвления аксона одного нейрона контактируют с телом клетки и дендритами другого нейрона.
2. Классификация нейронов. Нейроны различают по строению и функции. По строению (в зависимости от количества отходящих от тела клетки отростков) различают униполярные, псевдоуниполярные (оба отростка – аксон и дендрит – выходят из одного полюса клетки), биполярные (с двумя отростками) и мультиполярные (с множеством отростков) нейроны (рис. 2).
Истинно униполярные нейроны у человека обнаружены только в чувствительном ядре тройничного нерва на уровне среднего мозга.
По форме тела и характеру ветвления отростков выделяют звездчатые, пирамидные, веретеновидные, корзинчатые, зернистые и др. нейроны.
 |
По функциональным свойствам выделяют афферентные, чувствительные или центростремительные нейроны, несущие возбуждение от рецепторов в ЦНС, эфферентные, двигательные, мотонейроны, или центробежные, передающие возбуждение из ЦНС к иннервируемому органу, и вставочные, контактные, или промежуточные нейроны, соединяющие между собой афферентные и эфферентные пути.
Афферентные нейроны относятся к псевдоуниполярным, их тела лежат в спинномозговых ганглиях. Отходящий от тела клетки отросток Т-образно делится на две ветви, одна из которых идет в центральную нервную систему и выполняет функцию аксона, а другая подходит к рецепторам и представляет собой длинный дендрит.
Большинство эфферентных и вставочных нейронов относится к мультиполярным. Мультиполярные вставочные нейроны (самые многочисленные) в большом количестве располагаются в задних рогах спинного мозга, находятся и во всех других отделах ЦНС. Они могут быть и биполярными, как, например, нейроны сетчатки, имеющие короткий ветвящийся дендрит и длинный аксон. Промежуточные нейроны осуществляют связь между эффекторными и рецепторными нейронами.
Мотонейроны располагаются в основном в передних рогах спинного мозга, они передают информацию от нервного центра к исполнительным органам.
С точки зрения выделяемого медиатора нейроны бывают: холинергические, адренергические, дофаминергические, серотонинергические, глицинергические, ГАМК-ергические и т. д. Несмотря на то, что у нейрона множество разветвлений аксона, во всех его окончаниях выделяется один и тот же медиатор (правило Дейла).
По чувствительности к действию раздражителей нейроны делят на моно-, би-, и полисенсорные.
Моносенсорные нейроны располагаются чаще в первичных проекционных зонах коры и реагируют только на сигналы своей сенсорности. Например, значительная часть нейронов первичной зоны зрительной области коры большого мозга реагирует только на световое раздражение сетчатки глаза.
Моносенсорные нейроны подразделяют функционально по их чувствительности к разным качествам одного раздражителя. Так, например, отдельные нейроны слуховой зоны коры большого мозга могут реагировать на предъявления тона в 1000 Гц и не реагировать на тоны другой частоты. Они называются мономодальными. Нейроны, реагирующие на два разных тона, называются бимодальными, на три и более – полимодальными.
Бисенсорные нейроны. Чаще располагаются во вторичных зонах коры какого-либо анализатора и могут реагировать на сигналы как своей, так и другой сенсорности. Например, нейроны вторичной зоны зрительной области коры большого мозга реагируют на зрительные и слуховые раздражения.
Полисенсорные нейроны. Это чаще всего нейроны ассоциативных зон мозга; они способны реагировать на раздражение слуховой, зрительной, кожной и других рецептивных систем.
3. Нейроглия. Нейроглия – это вспомогательная и очень важная составная часть нервной ткани, связанная с нейронами генетически, морфологически и функционально. Эти клетки заполняют в мозге все пространство между нейронами, они мельче нейронов в 3 – 4 раза. Каждый нейрон окружен несколькими клетками нейроглии, которые равномерно распределены по всему мозгу и составляют около 40% его объёма. В коре больших полушарий их примерно в 5 раз больше, чем нервных клеток.
Нейроглия выполняет опорную, трофическую, защитную, изоляционную функции. Клетки нейроглии, возможно, принимают участие в обмене информацией внутри нейронного ансамбля, а также в условно-рефлекторной деятельности мозга и в процессах памяти. Глиальные клетки обладают способностью перемещаться в пространстве по направлению к наиболее активным нейронам. Это наблюдается при различных афферентных раздражениях и при мышечной нагрузке. Например, уже через 20 мин. плавания у крыс было обнаружено увеличение числа глиальных клеток вокруг мотонейронов переднего рога спинного мозга.
С возрастом количество нейронов в ЦНС уменьшается, а клеток нейроглии – увеличивается, так как последние, в отличие от нейронов, сохраняют способность к делению.
Нейроглия по своему происхождению подразделяется на макроглию и микроглию. Макроглия, как и нейроны, возникает из эктодермы, а микроглия развивается из мезодермы и является производным мезенхимы. В состав макроглии входят эпендима, астроглия и олигодендроглия.
Клетки эпендимы называются эпендимоцитами. Эпендимоциты образуют одинарный слой клеток ивыстилают полости нервной системы — спинномозговой канал, желудочки головного мозга, мозговой водопровод. Эпендимоциты имеют кубическую или цилиндрическую форму. На ранних стадиях развития у них есть реснички, обращенные в мозговые полости. Они способствуют проталкиванию цереброспинальной жидкости (ликвора). Позже реснички исчезают, сохраняясь лишь в некоторых участках, например в водопроводе.
Клетки эпендимы активно регулируют обмен веществами между мозгом и кровью, с одной стороны, и ликвором (цереброспинальной жидкостью) и кровью, с другой. Некоторые эпендимные клетки, танициты, имеют длинные цитоплазматические отростки, глубоко вдающиеся в ткань мозга. У таких эпендимоцитов в III желудочке (полости про межуточного мозга) отростки заканчиваются пластинчатым расширением на кровеносных капиллярах гипофиза. В этом случае эпендимоциты участвуют в транспорте веществ из ликвора в кровеносную сеть гипофиза

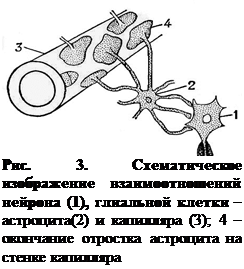 Астроглия (около 60% от общего числа клеток нейроглии) состоит из клеток – астроцитов, названных так потому, что от них радиально отходят многочисленные отростки. Астроглия выполняет опорную и трофическую функции (рис. 3). Отростки астроцитов образуют также пограничный слой на поверхности головного и спинного мозга – пограничную мембрану мягкой мозговой оболочки, прилегающую к базальной мембране. Это основной элемент гемато-энцефалического барьера; регулирует водно-солевой обмен нервной ткани. Астроциты образуют мостики между капиллярами и эпендимой, выстилающей полости желудочков мозга. Таким образом обеспечивается обмен между кровью и цереброспинальной жидкостью, т. е. астроциты выполняют транспортную функцию.
Астроглия (около 60% от общего числа клеток нейроглии) состоит из клеток – астроцитов, названных так потому, что от них радиально отходят многочисленные отростки. Астроглия выполняет опорную и трофическую функции (рис. 3). Отростки астроцитов образуют также пограничный слой на поверхности головного и спинного мозга – пограничную мембрану мягкой мозговой оболочки, прилегающую к базальной мембране. Это основной элемент гемато-энцефалического барьера; регулирует водно-солевой обмен нервной ткани. Астроциты образуют мостики между капиллярами и эпендимой, выстилающей полости желудочков мозга. Таким образом обеспечивается обмен между кровью и цереброспинальной жидкостью, т. е. астроциты выполняют транспортную функцию.
Олигоденроглия (около 25—30%) состоит из клеток, называемых олигодендроцитами, имеющих малое количество отростков. Отличаются высоким уровнем белкового и нуклеинового обмена; ответственны за транспорт веществ в нейроны. Они находятся в сером и белом веществе головного и спинного мозга, а также за пределами центральной нервной системы. В сером веществе центральных отделов нервной системы они окружают тела нейронов и их отростков, в белом веществе располагаются рядами или группами, а за пределами центральной нервной системы сопровождают отростки нервных клеток и известны под названием шванновских клеток, или нейролеммоцитов. Олигодендроциты синтезируют миелин – вещество белого цвета липоидной природы, которое обладает хорошими изоляционными свойствами (рис. 4). Изолируя отростки нервных клеток, олигоденроциты препятствуют рассеиванию нервного возбуждения. Кроме того, олигодендроциты принимают участие в питании нейронов и водном обмене мозга, поскольку всегда контактируют с кровеносными сосудами и, по некоторым данным, вместе с астроцитами участвуют в хранении следов возбуждения в нервной системе.
Некоторые клетки нейроглии выделяют вещества, влияющие на состояние возбудимости нервных клеток. Отмечено, что при разных психических состояниях изменяется секреция этих клеток.
Микроглия представлена клетками микроглии, или клетками Гортега, которые имеют небольшие древовидные отростки и отличаются малыми размерами. Основная их функция — защитная. Они являются фагоцитами нервной системы, за что их называют еще глиальными макрофагами. Количество этих клеток очень варьирует в зависимости от функционального состояния нервной системы. При различных экзо- и эндогенных вредных влияниях (травма, воспаление и т.п.) они резко увеличиваются в размерах, начинают делиться и устремляются в очаг поражения. Здесь микроглиоциты устраняют чужеродные клетки, например бактерии, и разного рода тканевые остатки (погибшие нервные клетки) и посторонние частицы путем фагоцитоза.
4. Нервные волокна, их виды, строение, свойства, возрастные изменения в связи с миелинизацией. Отросток нервной клетки, покрытый оболочками из глиальных клеток, называется нервным волокном. Центральную часть любого отростка нервной клетки (аксона или дендрита) называют осевым цилиндром. Осевой цилиндр покрыт оболочкой – аксолеммо й, под которой находится аксоплазма, содержащая нейрофибриллы и большое количество митохондрий.
В различных отделах нервной системы нервные волокна значительно различаются по своему строению, в связи с чем делятся на два типа: безмиелиновые и миелиновые. Миелиновая оболочка образуется шванновскими клетками путем их многократного накручивания вокруг осевого цилиндра и располагается не сплошной линией, а сегментами длиной 0,5 – 2 мм. Пространство между сегментами (1 – 2 мкм) называют перехватом Ранвье. В области перехватов Ранвье миелиновая оболочка отсутствует, и концы шванновских клеток плотно прилегают к аксолемме (рис. 4). Наружная мембрана шванновских клеток, покрывающая миелин, образует самую верхнюю оболочку нервного волокна, которую называют шванновской оболочкой, или неврилеммой. Безмиелиновые волокна отличаются тем, что в них не развивается миелиновая облочка и их осевые цилиндры покрыты только шванновскими клетками (шванновской оболочкой).
К миелинизированным относятся волокна соматической нервной системы и некоторые волокна вегетативной нервной системы. К безмиелиновым относится большинство волокон вегетативной нервной системы.
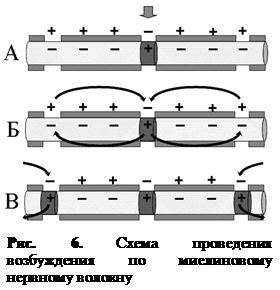
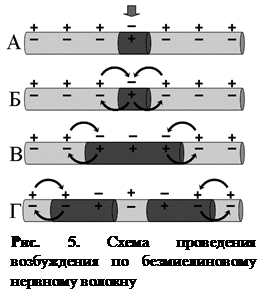
В безмякотных нервных волокнах возбуждение распространяется непрерывно вдоль всей мембраны, от возбужденного участка к невозбужденному, расположенному рядом, и сопровождается бóльшим расходом энергии, по сравнению с миелиновым волокном (рис. 5). Скорость проведения возбуждения относительно невелика. В отличие от этого в миелиновых волокнах возбуждение может распространяться только скачкообразно, «перепрыгивая» по перехватам Ранвье через участки волокна, покрытые миелином (рис. 6). Такое проведение называется сальтаторным. Возбуждение по мякотному нервному волокну проводится с очень большой скоростью. Расход энергии при этом невелик. Нервный импульс, возникший в одном перехвате Ранвье, способен вызывать возбуждение не только в лежащем рядом, но и в соседних 2–3 перехватах. Это создает гарантию проведения возбуждения по волокну, даже если 1 – 2 лежащих рядом перехвата повреждены.
 Нервные волокна делят на три группы, учитывая их строение и скорость проведения возбуждения: А, В и С. В группу «А» входят толстые миелиновые волокна, с диаметром 15 мкм и скоростью проведения возбуждения от 10 до 150 м/с. В группу «В» – тонкие миелиновые волокна диаметром 10 мкм, скоростью проведения возбуждения до 10 м/с; в группу «С» – тонкие безмиелиновые, диаметр 5 мкм, скорость поведения возбуждения до 1 м/с. В волокнах любого типа возбуждение распространяется без затухания (без декремента).
Нервные волокна делят на три группы, учитывая их строение и скорость проведения возбуждения: А, В и С. В группу «А» входят толстые миелиновые волокна, с диаметром 15 мкм и скоростью проведения возбуждения от 10 до 150 м/с. В группу «В» – тонкие миелиновые волокна диаметром 10 мкм, скоростью проведения возбуждения до 10 м/с; в группу «С» – тонкие безмиелиновые, диаметр 5 мкм, скорость поведения возбуждения до 1 м/с. В волокнах любого типа возбуждение распространяется без затухания (без декремента).
Основные свойства нервных волокон заключаются в следующем: высокая возбудимость и лабильность, низкий уровень обмена веществ, относительная неутомляемость, большая скорость проведения возбуждения. Проведение возбуждения по нервам подчиняется ряду законов: физиологической и анатомической целостности нерва, двустороннего проведения возбуждения от места возникновения, изолированного проведения возбуждения. Нервные импульсы распространяются по нервным волокнам без затухания или без декремента.
Миелинизация нервных волокон осуществляется в центробежном направлении, отступя несколько микрон от тела клетки к периферии нервного волокна. Отсутствие миелиновой оболочки резко ограничивает функциональные возможности нервного волокна, обусловливает диффузные, слабо координированные реакции.
По мере развития миелиновой оболочки возбудимость нервных волокон повышается. Раньше других начинают миелинизироваться периферические нервы, затем волокна спинного мозга, стволовой части головного мозга, мозжечка и позже – больших полушарий головного мозга. Этот процесс начинается на 4-м месяце внутриутробного периода и завершается в основном к 3 годам. Известно, что у новорожденного ребенка миелинизировано примерно 2/3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция внимания, он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций.
5. Строение синапса. Синапс – это морфофункциональное образование, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (от греческого слова «синапто» – контактировать). Термин и понятие «синапс», «синаптическая передача» был введен в физиологию Ч. Шеррингтоном в 1897 г.
Чем больше синапсов на нервной клетке, тем больше воспринимается различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия нервной клетки в разнообразных реакциях организма. На одном крупном мотонейроне насчитывают до 20 000 синапсов.
Количество синаптических контактов неодинаково на теле и отростках нейрона и очень вариабельно в различных отделах центральной нервной системы. Тело нейрона в среднем на 38% покрыто синапсами. Наибольшее число (до 50%) синапсов находится на дендритах. Особенно густо они покрывают средние части и окончания дендритных отростков, причем многие контакты расположены на специальных шипиках, которые еще больше увеличивают воспринимающую поверхность нейрона. Чем сложнее интегративная функция нейрона, тем большее развитие имеют аксодендритические синапсы (в первую очередь те, которые расположены на шипиках). Особенно они характерны для нейрональных связей пирамидных клеток в коре больших полушарий.
По способу передачи возбуждения синапсы подразделяют на электрические и химические и смешанные.
По местоположению синапсы подразделяются на нейро-мышечные (аксон нейрона контактирует с мышечной клеткой); нейро-секреторные (аксон нейрона контактирует с секреторной клеткой); нейро-нейрональные (аксон нейрона контактирует с другим нейроном)
По физиологическому эффекту синапсы подразделяют на возбуждающие и тормозные.
Синапс состоит из пресинаптического и постсинаптического отделов, между которыми имеется небольшое пространство, получившее название синаптической щели.
У электрического синапса клеточные мембраны соседних нейронов тесно прилегают друг к другу, так что между ними остается только очень узкая щель шириной 2 – 5 нм. Обе мембраны соединяются посредством поперечных каналов, образованных особыми белками-коннексонами. Эти каналы не сообщаются с внеклеточным пространством и отсутствуют в других участках мембраны. Они имеют низкую утечку во внешнюю среду и передают нервные импульсы с использованием того же механизма, что и при передаче по нервному волокну.
Электрические синапсы находятся там, где нужна быстрая передача, они биполярны, симметричны, сравнительно мало утомляемы, устойчивы к изменениям внешней и внутренней среды, проводят только возбуждение и возбуждением могут охватывать сразу несколько нейронов. У человека, также как и у млекопитающих, электрические синапсы находятся в нескольких областях мозга: в мезэнцефальном ядре тройничного нерва, в вестибулярном ядре Дейтерса, в нижней оливе, между некоторыми клетками в сетчатке.
Химические синапсы очень специфические, не симметричные, односторонние, между мембранами имеется щель. На прохождение щели уходит время. В отличие от электрических проводят как возбуждение, так и торможение. Химические синапсы обладают высокой утомляемостью. Это объясняется истощением запасов медиатора.
 Синапсы, образованные аксоном и телом (сомой) клетки, называют аксо-соматическими, аксоном и дендритом – аксо-дендритическими, контакты между аксонами двух нейронов получили название аксо-аксональных синапсов (рис. 7).
Синапсы, образованные аксоном и телом (сомой) клетки, называют аксо-соматическими, аксоном и дендритом – аксо-дендритическими, контакты между аксонами двух нейронов получили название аксо-аксональных синапсов (рис. 7).
Пресинаптический отдел химического синапса представлен конечной веточкой аксона, которая на расстоянии 200 – 300 мкм от контакта теряет миелиновую оболочку и на конце утолщается (рис. 8). Это утолщение называется синаптической бляшкой. В пресинаптическом отделе синапса содержится большое количество митохондрий и пузырьков (везикул) округлой или овальной формы размером от 0,02 до 0,05 мкм. В везикулах содержится вещество, обеспечивающее передачу возбуждения с одного нейрона на другой, которое называют медиатором. Везикулы концентрируются вдоль поверхности пресинаптической мембраны, находящейся против синаптической щели, ширина которой равна 0,0012 – 0,03 мкм.
Постсинаптический отдел синапса образуется мембраной сомы нейрона или его отростков, а в концевой пластинке (синапс между окончанием аксона и мышцей) – мембраной мышечного волокна. Постсинаптическая мембрана несет на себе белковые хеморецепторы к биологически активным веществам. Важная особенность рецепторов постсинаптической мембраны – их химическая специфичность, т. е. способность вступать в биохимическое взаимодействие только с определенным видом медиатора.
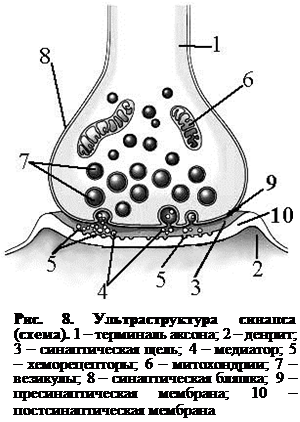 Пресинаптическая и постсинаптичес-кая мембраны имеют специфические особенности строения, связанные с передачей возбуждения: они несколько утолщены (их диаметр около 0,005 мкм). Длина этих участков составляет 150 – 450 мкм. Утолщения могут быть сплошными и прерывистыми. Постсинаптическая мембрана у некоторых синапсов может быть складчатой, что увеличивает поверхность соприкосновения ее с медиатором.
Пресинаптическая и постсинаптичес-кая мембраны имеют специфические особенности строения, связанные с передачей возбуждения: они несколько утолщены (их диаметр около 0,005 мкм). Длина этих участков составляет 150 – 450 мкм. Утолщения могут быть сплошными и прерывистыми. Постсинаптическая мембрана у некоторых синапсов может быть складчатой, что увеличивает поверхность соприкосновения ее с медиатором.
Проходящий по терминали аксона нервный импульс достигает пресинаптической мембраны, что приводит к её деполяризации и открытию кальциевых пор (каналов). Кальций мигрирует через эти каналы внутрь аксонной терминали и способствует слиянию везикул с пресинаптической мембраной и опорожнению медиатора (экзоцитозу) в синаптическую щель. Количество медитора, которое освобождается из одного синаптического пузырька, называется квантом медиатора. Под действием одного нервного импульса в синаптическую щель освобождается 100 – 200 квантов медиатора.
Выделившийся медиатор диффундирует к постсинаптической мембране. Этот процесс требует около 0,5 мс и вносит существенный вклад в задержку передачи. Взаимодействие медиатора со своими рецепторами вызывает возбуждение или торможение нейрона, сокращение мышечной клетки, образование и выделение гормонов секреторными клетками. Передача возбуждения происходит только в одном направлении: от синаптического окончания к постсинаптической мембране.
В синапсах ЦНС, как и в концевой пластинке, в состоянии относительного покоя отмечено спонтанное разрушение везикул и выход квантов медиатора в синаптическую щель. Этого недостаточно для возникновения возбуждения в постсинаптической мембране, однако синапс поддерживается в состоянии готовности к передаче.
Инактивация (полная потеря активности) медиатора осуществляется путем гидролитического расщепления с помощью соответствующих ферментов. Ацетилхолин (АХ) расщепляется ферментом холинэстеразой, норадреналин (НА) и адреналин – моноаминоксидазой (МАО) и катехолоксиметилтрансферазой (КОМТ). Продукты расщепления медиатора снова поступают в кровь и циркулируют, как его предшественники.
Другой путь удаления медиатора из синаптической щели – «обратный захват» его пресинаптическими окончаниями (пиноцитоз) и обратный аксонный транспорт, особенно выраженный для катехоламинов. Это создает возможность их быстрого поступления в синаптическую щель под влиянием нового нервного импульса.
Для обеспечения нормальной работы химического синапса требуется поддержание необходимого количества везикул и медиатора. Некоторые медиаторы (например, ацетилхолин) синтезируются в цитоплазме синаптического окончания, и там же молекулы медиатора депонируются в синаптических пузырьках. Ферменты, необходимые для синтеза медиатора, образуются в теле нейрона и доставляются в синаптическое окончание путем медленного (1–3 мм/сут) аксонного транспорта. Другие медиаторы (пептиды и др.) синтезируются и упаковываются в везикулы в теле нейрона, готовые синаптические пузырьки доставляются в синаптичекую бляшку за счет быстрого (400 мм/сут) аксонного транспорта. Синтез медиатора и образование синаптических пузырьков осуществляется непрерывно.
В ЦНС выделяют два типа нейронов: возбуждающие и тормозящие. Возбуждающие нейроны выделяютв своих синапсах такие медиаторы, как ацетилхолин (АХ), норадреналин, серотонин, кислые аминокислоты. Тормозящие нейроны вызывают торможение за счет медиаторов, таких как глицин, β-аланин, γ-аминомасляниая кислота (ГАМК), дофамин. В то же время установлено, что один и тот же медиатор, например АХ, может выделяться как возбуждающими, так и тормозными синапсами, и, соответственно, вызывать как возбуждение, так и торможение. Поэтому можно говорить об амбивалентности медиаторов.
Каждый синаптический контакт на теле или отростках нейрона способствует развитию местных процессов возбуждения или торможения. На мембране нейрона происходит их алгебраическое суммирование. При одновременной активации нескольких возбуждающих синапсов их общий эффект представляет собой сумму отдельных местных (локальных) процессов возбуждения. При одновременном возникновении двух различных синаптических влияний (возбуждающих и тормозящих) происходит взаимное вычитание их эффектов. В конечном итоге реакция нервной клетки определяется суммой всех синаптических влияний.
Активная деятельность нервных клеток способствует улучшению синаптического взаимодействия между ними: росту площади синаптических контактов, ускорению проведения возбуждения через синапсы и т. д. Это наблюдается, например, при двигательной деятельности. Как показали эксперименты на животных, ежедневная принудительная ходьба их по 1 часу приводит к значительной активации синапсов в соответствующих рефлекторных дугах спинного мозга.
Синапсы, являются самым чувствительным местом нервной системы. Именно синаптические контакты между нейронами первыми реагируют на различные повреждающие воздействия. При травмах нервной ткани, отеках, нарушениях кровообращения, рентгеновских облучениях, отравлениях и других неблагоприятных воздействиях в синапсах сначала возникают обратимые изменения - сильное набухание (увеличение их размеров в несколько раз) и деформация, а затем происходят необратимые изменения синапсов - их разрушение, в результате чего полностью нарушается нервная деятельность.
6. Рефлекс и рефлекторная дуга. Ответную реакцию организма на раздражение рецепторов, осуществляющуюся с участием центральной нервной системы, называют рефлексом. Раздражителем, вызывающим рефлекторную реакцию, может быть любое изменение внешней и внутренней среды организма.
Время от момента нанесения раздражения до появления ответной реакции называют временем рефлекса. Время рефлекса зависит от количества нейронов (синапсов), входящих в рефлекторную дугу, а также от интенсивности раздражения: чем больше сила раздражения, тем меньше время рефлекса, и, наоборот, чем меньше сила раздражения, тем больше время рефлекса.
Путь, по которому нервный импульс распространяется от рецептора к эффектору (т. е. осуществляется рефлекс), называют рефлекторной дугой. Рефлекторная дуга состоит из пяти основных звеньев: рецептора, афферентного пути, центральной нервной системы, эфферентного пути и эффектора.
Простейшие рефлекторные дуги состоят из двух или трех нейронов (рис. 4).
|

|

 |
Наиболее простая связь между нейронами имеет место в двухнейронной рефлекторной дуге. В ней аксон афферентного нейрона непосредственно контактирует с телом клетки и дендритами мотонейрона в передних рогах спинного мозга. Особенностью двухнейронной дуги является то, что и рецептор и эффектор рефлексов, осуществляющихся по этому типу, лежат в одном и том же органе. К двухнейронным относят сухожильные рефлексы (ахиллов, коленный и др.).
В трехнейронную рефлекторную дугу входят афферентный, вставочный и эфферентный нейроны. Дендрит афферентного нейрона начинается от рецептора, тело афферентного нейрона лежит в спинномозговом ганглии. Аксон афферентного нейрона от ганглия идет в составе задних корешков в центральную нервную систему и в задних рогах спинного мозга контактирует со вставочным нейроном. Аксон вставочного нейрона передает возбуждение в передние рога спинного мозга, к мотонейрону, аксон которого несет возбуждение к иннервируемому им органу.
В большинстве рефлекторных актов связи между нейронами сложнее, чем это представлено в трехнейронной рефлекторной дуге, и вставочных нейронов в их дугах может быть много.
Рефлекторные дуги, в которых возбуждение проходит через один синапс, называют моносинаптическими, а те, в которых возбуждение последовательно проходит более чем через один синапс, – полисинаптическими.
Рефлекторный акт не заканчивается ответной реакцией организма на раздражение. В работающем органе раздражаются рецепторы, и возникающее в них возбуждение по афферентным волокнам возвращается в центральную нервную систему. Каждый орган, будь это скелетные мышцы, осуществляющие движение, или ритмически сокращающаяся сердечная мышца, или мышцы постоянно пульсирующих сосудов, дыхательные мышцы и т. д., благодаря обратной импульсации «сообщает» нервным центрам о своем состоянии. В ответ на полученный афферентный сигнал нервный центр в каждый данный момент вносит свои поправки в осуществляющийся рефлекторный акт. Поэтому принято говорить не о рефлекторной дуге, а о рефлекторном кольце.
7. Классификация рефлексов. Классифицируя многообразные рефлексы организма человека и животного, учитывают их различные признаки и проявления.
Все рефлексы по происхождению делят на безусловные – врожденные – видовые рефлексы и условные – приобретенные в процессе индивидуальной жизни животного или человека, выработанные при определенных условиях.
На основании биологического значения тех или иных рефлексов для организма их делят на оборонительные, или защитные, направленные на удаление от раздражителя, пищевые, обеспечивающие добывание, потребление и переваривание пищи, половые, связанные с продолжением рода, ориентировочные, или исследовательские, проявляющиеся в повороте тела и движении в сторону нового раздражителя, позно-тонические, или рефлексы положения тела в пространстве, и локомоторные – перемещения тела в пространстве.
В зависимости от расположения рецепторов рефлекторной дуги, различают экстероцептивные рефлексы, возникающие в ответ на раздражение рецепторов поверхности тела, проприоцептивные – на раздражение рецепторов мышц, сухожилий и суставов, висцероцептивные – на раздражение рецепторов внутренних органов.
Классифицируют рефлексы и по органам, отвечающим на раздражение. При этом говорят о сердечных, дыхательных, сосудистых и других рефлексах.
Рефлексы различают также по характеру ответных реакций: секреторные, выражающиеся в выделении секрета, выработанного железой, трофические, связанные с изменением обмена веществ, и двигательные, или моторные, характеризующиеся сократительной деятельностью поперечнополосатых и гладких мышц. Последние составляют самую разнообразную группу рефлексов. При раздражении кожи возникают сгибательные, потирательные рефлексы, рефлексы почесывания и др. Раздражением кожи губ у ребенка вызывается сосательный рефлекс, роговицы глаза – мигательный. Сужение зрачка при действии света и его расширение в темноте представляют собой зрачковый рефлекс.
Двигательные проприоцептивные рефлексы возникают при раздражении рецепторов мышц и сухожилий. Так, при ударе по сухожилию четырехглавой мышцы бедра в результате ее растяжения возникает рефлекторное разгибание ноги в колене – коленный рефлекс, при ударе по ахиллову сухожилию – ахиллов рефлекс.
Рефлексы, заключающиеся в сужении и расширении сосудов, называются сосудодвигательными.
Двигательные рефлексы, возникающие при раздражении рецепторов гладкой мускулатуры внутренних органов, носят название висцеромоторных. С ними связаны движения желудка, кишечника, мочевого пузыря, мочеточников и др.
Все описанные выше рефлексы в зависимости от того, какие отделы центральной нервной системы участвуют в их осуществлении, делят на спинальные (осуществляющиеся с участием нейронов спинного мозга), бульбарные (с участием нейронов продолговатого мозга), мезенцефальные (с участием среднего мозга), диэнцефальные (с участием промежуточного мозга) и кортикальные (с участием нейронов коры больших полушарий головного мозга).
Примером спинальных рефлексов могут быть сгибательный, возникающий при пощипывании лапки лягушки пинцетом, потирательный – при раздражении кожи лягушки бумажкой, смоченной серной кислотой, и др. К ним же относятся рефлексы сухожильного типа.
Сосательный, мигательный рефлексы осуществляются с участием продолговатого, зрачковый – среднего мозга. Регуляция любой функции осуществляется с участием разных отделов центральной нервной системы. Поэтому классификация рефлексов по отделам мозга, участвующим в их осуществлении, относительна. Речь идет только о преимущественном значении нейронов того или иного отдела центральной нервной системы.
8. Классификация и строение рецепторов. Живые организмы не могут существовать, не получая информации из внешней и внутренней среды. Восприятие энергии раздражителя и преобразование ее в нервный импульс осуществляют рецепторы.
Рецептор представляет собой окончание дендрита афферентного аксона или комплекс нервной и эпителиальной клеток, специально приспособленный для восприятия определенного типа раздражителей. Рецепторы являются начальными звеньями любой рефлекторной дуги, а также участвуют в оценке параметров полезного приспособительного результата в функциональных системах организма.
Дендрит каждого афферентного нейрона выходит на периферию и разветвляется на многочисленные тончайшие веточки – терминали. На конце каждой терминали находится утолщение – рецепторная бляшка.
По строению рецепторы подразделяют на первичные (первичночувствующие) и вторичные (вторичночувствующие) (рис. 10).
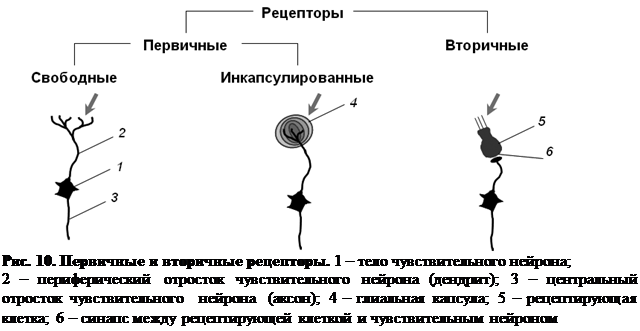 |
К первичным относят такие рецепторы, у которых действие раздражителя воспринимается непосредственно рецепторной бляшкой, которая может быть либо свободной, т. е. не иметь дополнительных образований, либо инкапсулированной. В этом случае рецепторная бляшка заключена в особую капсулу, осуществляющую первичное преобразование энергии раздражителя. К первичночувствующим рецепторам относятся: обонятельные, тактильные рецепторы и мышечные веретена.
К вторичным относят такие рецепторы, у которых действие раздражителя воспринимается специализированной рецептирующей клеткой не нервного происхождения. Возбуждение, возникшее в рецептирующей клетке, передается через синапс на чувствительный нейрон, который возбуждается не непосредственно стимулом, а опосредовано (вторично) потенциалом рецептирующей клетки. К вторичночувствующим рецепторам относятся: рецепторы слуха, зрения, вкуса, вестибулярные рецепторы.
По расположению рецепторы подразделяют на: экстерорецепторы – воспринимают раздражители из внешней среды организма; интерорецепторы – воспринимают раздражители из внутренней среды организма. Они в свою очередь подразделяются на проприорецепторы – специализированные рецепторы опорно-двигательной системы (мышц, сухожилий, суставов), вестибулорецепторы – воспринимают положение тела в пространстве и висцерорецепторы – получают сигналы от внутренних органов.
По разнообразию воспринимаемых раздражителей рецепторы подразделяют на: мономодальные – приспособленные для восприятия только одного вида раздражителя, и полимодальные – приспособленные для восприятия различных раздражителей.
По модальности рецепторы подразделяют на: хеморецепторы – воспринимают действие химических веществ; фоторецепторы – воспринимают световые раздражители; механорецепторы – воспринимают давление, вибрацию, перемещение, степень растяжения; терморецепторы – чувствительны к изменениям температуры; ноцицепторы – воспринимают болевое раздражение.
 По способности воспринимать раздражитель, находящийся на расстоянии от рецептора или при непосредственном контакте с ним, различают дискантные и контактные рецепторы
По способности воспринимать раздражитель, находящийся на расстоянии от рецептора или при непосредственном контакте с ним, различают дискантные и контактные рецепторы
Все рецепторы обладают следующими свойствами.
1. Специфичность – способность воспринимать определенный, т. е. адекватный данному рецептору, раздражитель. Эта способность рецепторов сформировалась в процессе эволюции.
2. Высокая чувствительность – способность реагировать на очень малые по интенсивности параметры адекватного раздражителя. Например, для возбуждения фоторецепторов сетчатки глаза достаточно нескольких, а иногда и одного, квантов света. Обонятельные рецепторы информируют организм о появлении в атмосфере единичных молекул пахучих веществ.
3. Способность к ритмической генерации импульсов возбуждения в ответ на однократное действие раздражителя.
4. Возбуждение рецепторов наступает после некоторого латентного периода и имеет последействие.
5. Способность к адаптации – т. е. способность приспосабливаться ("привыкать") к постоянно действующему стимулу. Адаптация может выражаться в снижении активности рецептора и частоты генерации импульсов возбуждения, вплоть до полного его прекращения. В зависимости от скорости адаптации различают: быстроадаптирующиеся (тактильные), медленноадаптирующиеся (терморецепторы), неадаптирующиеся (болевые, вестибулярные и проприорецепторы).
Выделяют несколько видов адаптации: изменение возбудимости рецептора в сторону снижения – десенсибилизация; изменение возбудимости в сторону повышения – сенсибилизация.
При действии сильных раздражителей, порог чувствительности повышается. Например: если человек вышел из тепла сразу в холод – то человеку вначале холодно, но затем рецепторы повышают свой порог и человеку становится тепло.
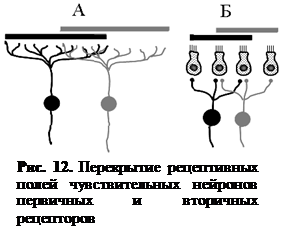 Рецептивное поле. Рецептивным полем нейрона называют множество рецепторов, функционально связанных с этим нейроном. Рецептивное поле нейрона представляет собой динамическое образование – один и тот же нейрон в различные моменты времени может оказаться функционально связанным с различным количеством рецепторов. Максимальная величина рецептивного поля какого-либо нейрона соответствует количеству рецепторов, которые связаны с эти нейроном морфологически, а минимальная величина может ограничиваться всего одним рецептором (рис. 11).
Рецептивное поле. Рецептивным полем нейрона называют множество рецепторов, функционально связанных с этим нейроном. Рецептивное поле нейрона представляет собой динамическое образование – один и тот же нейрон в различные моменты времени может оказаться функционально связанным с различным количеством рецепторов. Максимальная величина рецептивного поля какого-либо нейрона соответствует количеству рецепторов, которые связаны с эти нейроном морфологически, а минимальная величина может ограничиваться всего одним рецептором (рис. 11).
У первичных рецепторов зоны ветвления периферических отростков чувствительных нейронов могут перекрывать друг друга (рис. 12, А). У вторичных рецепторов одна рецептирующая клетка может контактировать с несколькими чувствительными нейронами, т. е. может входить в состав рецептивных полей различных нейронов (рис. 12, Б).
Рецептивным полем рефлекса называют совокупность рецепторов, раздражение которых вызывает данный тип рефлекса.
|
|
Дата добавления: 2015-05-26; Просмотров: 3487; Нарушение авторских прав?; Мы поможем в написании вашей работы!