КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Индикаторы силы нервной системы по отношению к торможению 1 страница
|
|
|
|
Как и всякая работа по изучению свойств нервной системы, работа по изучению природы и проявлений силы относительно торможения должна была начаться с изыскания методик определения этого параметра нервной деятельности. Прецедент в этом отношении был дан И.П. Павловым, который в числе индикаторов «абсолютной силы тормозного процесса» называл такую пробу, как продление действия дифференцировочного раздражителя.
Известно, что в работе с животными этот прием использовался неоднократно и с успехом; собственно, он один дает эффективную количественную меру силы нервной системы относительно торможения у собак при работе по слюнной методике: количество капель слюны, выделившихся во время действия продленного дифференцировочного раздражителя, служит показателем «растормаживания», т. е. перехода за предел тормозной функции, и, следовательно, показателем силы – слабости нервной системы по отношению к действию тормозного процесса.
В той же роли может быть использован такой прием, как многократное повторение дифференцировочного раздражителя, судя по некоторым данным, коррелирующее с удлинением дифференцировки (Викт. К. Федоров, 1961). Кроме того, для целей дифференциальной диагностики могут быть использованы данные Ф.П. Майорова (1962), Е.Г. Гусевой (1959, 1961), Л.О. Зевальда (1964) и других, показывающие, что предел выносливости к действию тормозного стимула может быть достигнут и при усилении физической интенсивности последнего.
Что касается фармакологических проб силы нервной системы по отношению к торможению, то в настоящее время таковых, видимо, не существует: В.К. Красуский (1953) и Т.Ф. Комарова (1960) показали, что испытание бромом для этой цели не подходит, а другие фармакологические индикаторы этого параметра, насколько нам известно, еще не разработаны.
|
|
|
Первым опытом разработки методики для определения силы нервной системы по отношению к торможению у человека в лаборатории Б.М. Теплова явилось исследование В.И. Рождественской, в котором в рамках методики фотохимических условных реакций измерялось влияние удлинения и многократного повторения дифференцировочного раздражителя на абсолютную световую чувствительности (1963а).
Опыты В.И. Рождественской были проведены в двух экспериментальных сериях: в одной из них испытывалось действие удлинения дифференцировки, в другой – влияние 10‑кратного предъявления дифференцировочного стимула. В первой серии проводилось по 2 – 3 опыта с каждой из двух длительностей дифференцировки: 20 и 30 с. Во второй серии длительность стимула была обычной, составляя 10 с, состояла серия из 2 – 3 опытов. Чувствительность измерялась после каждого предъявления дифференцировочного сигнала. Для контроля данных второй серии, в которой дифференцировка предъявлялась (и чувствительность измерялась) каждую минуту и в результатах которой можно было поэтому подозревать влияние сенсорного утомления, были проведены дополнительные опыты с измерением динамики порогов без предъявления дифференцировки.
В табл. 23, заимствуемой из статьи В.И. Рождественской (1963а), представлены результаты первой серии экспериментов.
Как видно из табл. 23, 2‑й 3‑кратное удлинение дифференцировки вызывает эффект растормаживания, выражающийся в таком понижении чувствительности, которое обычно вызывается положительным условным сигналом. Явное растормаживание, однако, наблюдается не у всех испытуемых, а только у Ш., Р. и И., причем у двух последних оно заметно уже при 20‑секундной длительности дифференцировки. У двух других испытуемых – Е. и Э. – растормаживание находится в пределах ошибки измерений, а у испытуемого Б. нет даже намека на это явление.
|
|
|
Данные второй серии для двух испытуемых иллюстрируются графиками, представленными на рис. 40 и 41. На рис. 40 показаны результаты испытуемого Б., который в первой серии не обнаружил никакой тенденции к рас-тормаживанию при удлинении дифференцировки, а на рис. 41 – результаты испытуемого И., показавшего наибольшее растормаживание. Разница между графиками очевидна. Она заключается в том, что у испытуемого Б. 10‑кратное повторение дифференцировки в каждом из трех опытов неизменно приводит к повышению чувствительности (реакция «обратного знака»), а у испытуемого И. в точности такой же эксперимент имеет каждый раз результатом понижение чувствительности, т. е. растормаживание дифференцировки. Данные испытуемых Е и Э. близки к данным испытуемого Б., а данные испытуемых Р. и Ш. – к данным испытуемого И.
Таблица 23
Изменение световой чувствительности (в %) после действия условного и дифференцировочного раздражителей разной длительности по сравнению с фоновым уровнем чувствительности (в. и. Рождественская, 1963а)
| Испытуемые | Условный раздражитель 10 с | Дифференцировочный раздражитель, с | ||
| Б. | –19 | +5 | +2 | |
| Е. | –24 | +1 | –4 | –4 |
| Э. | –17 | +4 | +13 | -8 |
| Ш. | –23 | +1 | -9 | -22 |
| Р. | -23 | +2 | -21 | -20 |
| и. | –21 | –20 | –37 |
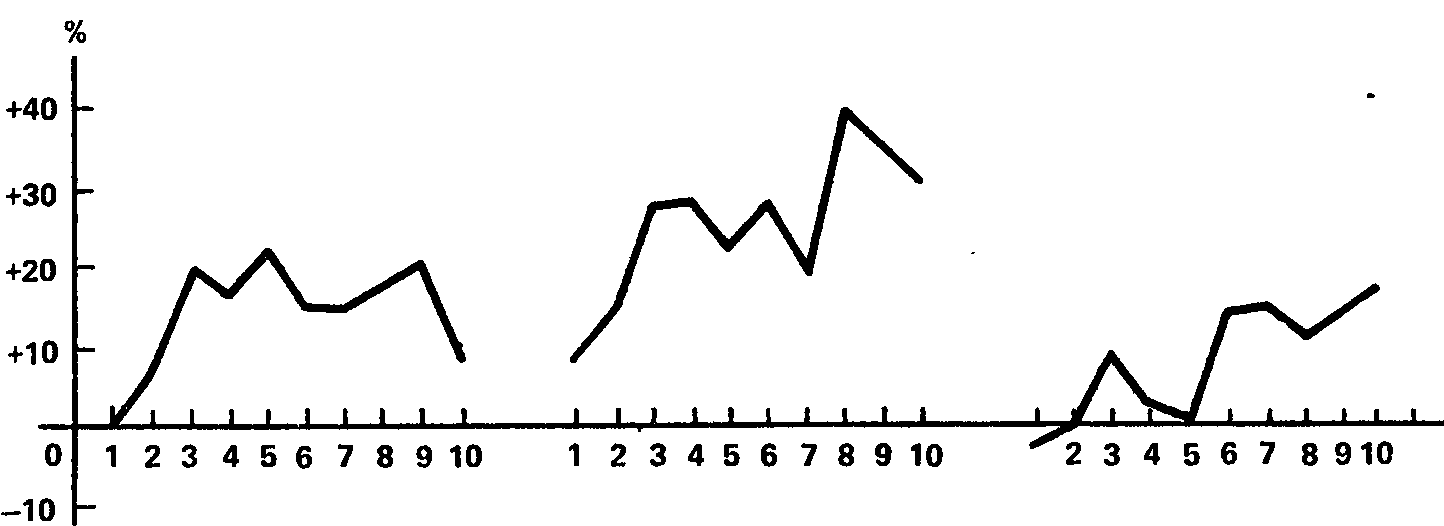
Рис. 40. Тенденция к росту световой чувствительности при 10‑кратном повторении дифференцировочного раздражения с интервалом 1 мин у испытуемого Б в трех различных опытах.
Ось абсцисс – порядковый номер предъявлений дифференцировки; ось ординат – изменение чувствительности (в %) по отношению к «фону» (В.И. Рождественская, 1963а).
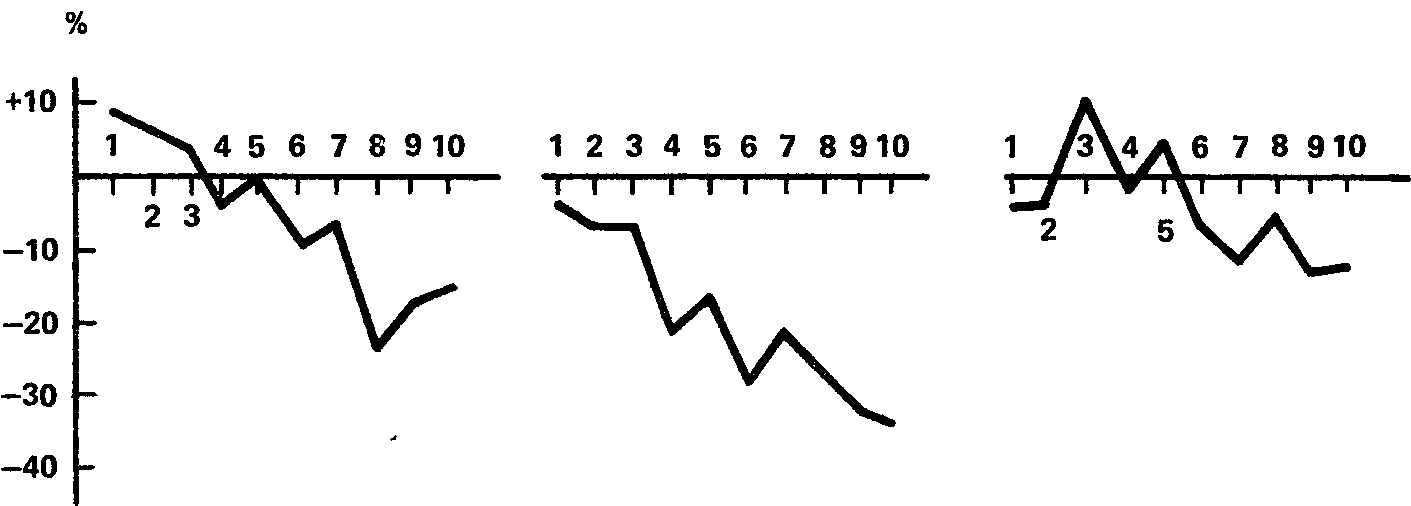
Рис. 41. Тенденция к падению световой чувствительности при 10‑кратном пов- – торении дифференцировки с интервалом 1 мин у испытуемого И. в трех различных опытах.
Обозначения те же, что на рис. 40 (В.И. Рождественская, 1963а).
Таким образом, и во второй серии экспериментов у части испытуемых дифференцировочный раздражитель, несмотря на форсирование тормозного процесса, сохраняет свое значение тормозного стимула, в то время как у другой части испытуемых аналогичный эксперимент приводит к тому, что дифференцировочный сигнал теряет способность вызывать тормозной эффект и действует подобно положительному раздражителю. При этом существенно, что результаты обеих серий, обоих способов усиления тормозного процесса в очаге дифференцировочного раздражителя в основном совпадают. Это дает право считать, что в основе индивидуальных вариаций, полученных при использовании двух различных приемов, лежит одно и то же свойство нервной системы. Исходя из функционального значения этих приемов, можно, вслед за В.И. Рождественской, с большой долей уверенности полагать, что этим свойством является сила (выносливость, работоспособность) нервной системы по отношению к тормозному процессу.
|
|
|
Таким образом, показателем собственно силы нервных клеток по отношению к торможению в методике фотохимических условных реакций является либо отсутствие изменений световой чувствительности при усилении очага торможения, либо повышение чувствительности – реакция «обратного знака». Признаком слабости нервной системы по отношению к торможению служит в аналогичных условиях понижение чувствительности при действии дифференцировки в результате ее растормаживания.
Насколько перспективен «фотохимический» способ оценки силы нервной системы по отношению к торможению? Пригоден ли он для широкого употребления хотя бы в лабораторной практике? С большим сожалением на этот вопрос, видимо, придется ответить отрицательно. Хотя процедура получения требуемого количественного показателя сама по себе достаточно коротка и к тому же, по всей видимости, дает достаточно надежные (в статистическом смысле) результаты, значительные препятствия на пути ее широкого применения воздвигаются несоразмерно большой длительностью и трудоемкостью предварительной работы по выработке стойких положительных и дифференцировочных условных реакций.
Однако изложенная работа В.И. Рождественской имеет немаловажное принципиальное значение. Если при помощи какой-то одной рефлекторной методики в принципе возможно получение у человека индикаторов силы нервной системы относительно торможения, то, очевидно, это возможно при использовании и других рефлекторных реакций. Попытка использовать с этой целью кожно-гальваническую методику была недавно предпринята Л.Б. Ермолаевой-Томиной (1963), которая ввела в сопоставление целого ряда ориентировочных и условнорефлекторных показателей и такие пробы, как удлинение и 15‑кратное повторение дифференцировки.
|
|
|
Оказалось, что растормаживание дифференцировки при ее удлинении фактически является функцией баланса нервных процессов и происходит исключительно у лиц с преобладанием возбуждения. Несомненно, в этом сказывается эмоционально-ориентировочная природа кожно-гальванической реакции: эффект ориентировки при изменении ситуации (удлинение сигнала) и эффект растормаживания выражаются в одном и том же колебании потенциала. В связи с этим судить о причине возникновения КГР при удлинении дифференцировки довольно трудно: не ясно, обусловлено ли оно действительным перенапряжением тормозного процесса или просто новизной в характере предъявления стимула.
Что же касается многократного повторения дифференцировочного раздражителя, то его результаты с показателями баланса не коррелировали. Это обстоятельство может служить доводом в пользу предположения о том, что повторение дифференцировки измеряет качество работы нервной системы, отличное от баланса по динамичности, возможно, силу по отношению к торможению. Однако доказательств этого мы до сих пор не имеем.
Таким образом, возможности кожно-гальванической методики в аспекте разработки индикаторов силы нервной системы относительно торможения (как, впрочем, и возбуждения) представляются довольно ограниченными; не исключено, однако, что эти возможности еще не исчерпаны, как не исчерпаны еще в этом аспекте возможности целого ряда других психофизиологических методик и индикаторов. Задача изыскания адекватных методических приемов для определения у человека силы нервной системы по отношению к торможению представляет очень большую важность, и решения этой задачи следует искать на самых различных методических путях.
Глава 8. Сила нервной системы и абсолютная чувствительность
Создание эффективных и надежных методов определения силы нервной системы[30] позволило осуществить разносторонние исследования природы и проявлений этого важного параметра нервной деятельности. В целом ряде экспериментальных работ, проведенных в лаборатории Б.М. Теплова, было показано существование комплекса разнообразных качеств нервной деятельности, группирующихся вокруг параметра силы и составляющих в совокупности синдром проявлений этого свойства нервной системы.
Комплексность проявлений является необходимым формальным признаком свойства нервной системы как стационарного параметра ее организации. «Нельзя представить себе такого основного свойства нервной системы, которое имело бы только одно проявление. Это, будет частная особенность нервной системы, но никак не основное свойство ее» (Б.М. Теплов, 1963, с. 8). Что касается силы нервной системы, то существование вокруг нее такого комплекса проявлений и зависимостей в настоящее время является несомненным, и одним из наиболее существенных атрибутов этого свойства является его неразрывная внутренняя связь с абсолютными порогами ощущений.
Получившая ныне широкую известность гипотеза Б.М. Теплова о зависимости между чувствительностью, реактивностью нервной системы и ее силой была впервые выдвинута (1955) в форме чисто теоретического умозаключения, выведенного на основе анализа некоторых высказываний И.П. Павлова о функциональных качествах корковых клеток, анализа эффекта применяемых методов повышения возбудимости, а также некоторых наблюдений различных авторов над особенностями поведения животных слабого типа.
Б.М. Теплов обратил специальное внимание на те высказывания И.П. Павлова, в которых речь шла о причинной зависимости между уровнем «высшей реактивности» корковой клетки и пределом ее работоспособности. И.П. Павлов полагал, что именно исключительной реактивностью и, как следствие этого, стремительной функциональной разрушаемостью клетки коры отличаются от других клеток нервной системы. «Не следует ли аналогичным образом понимать и различия между корковыми клетками слабой и сильной нервной систем?» – спрашивает он. Б.М. Теплов (1955, с. 6) отвечает на этот вопрос положительно. Таким образом, было дано гипотетическое объяснение специфической особенности слабой нервной системы, заключающейся в низком пределе ее работоспособности и склонности к быстрому развитию запредельного торможения; эти качества были поставлены в связь с высокой реактивностью, возбудимостью, чувствительностью нервных клеток слабой нервной системы.
Нужно сказать, что в момент выдвижения обсуждаемой гипотезы понятия реактивности, возбудимости и чувствительности принимались как синонимические, рядоположные. Впоследствии, однако, возникла необходимость в их уточнении и определенном разграничении, поскольку (чтобы не вводить новые термины) каждое из них полезнее использовать для обозначения, по крайней мере, частично специфического круга явлений. Это особенно относится к понятию реактивности в сопоставлении с двумя другими понятиями.
Если понятиями чувствительности и возбудимости подчеркивается содержание, относящееся к порогу реакции, к минимальной величине стимула, вызывающей состояние возбуждения, то в понятии реактивности, видимо, основным является момент величины самой реакции, на основе которой судят о наличии раздражения. Но по величине реакции не во всех случаях можно судить о величине раздражения. Вмешательство некоторых факторов, относящихся как к общим, так и к индивидуальным особенностям работы нервной системы, может привести к тому, что характеристика по реактивности не будет совпадать с характеристикой по чувствительности, возбудимости; так, оказывается, что меньший по интенсивности (пороговый) сигнал вызывает большую по величине реакцию некоторых вегетативных компонентов ориентировочного рефлекса, чем сверхпороговый раздражитель (О.С. Виноградова, Е.Н. Соколов, 1955), и может также оказаться, что менее чувствительная система явится более реактивной, т. е. даст большую величину реакции, чем более чувствительная (равно как и наоборот). Отсюда следует, что при характеристике функции порога предпочтительнее пользоваться понятиями чувствительности или возбудимости, чем понятием реактивности.
Что касается различия между понятиями чувствительности и возбудимости, то оно является более частным и сводится к тому, что первое понятие обычно применяется при определении порогов ощущений и, стало быть, возможно только по отношению к функции организма как целого, а второе более употребляется при измерении пороговых характеристик возбудимых тканей. Принципиальной же разницы между понятиями чувствительности и возбудимости, очевидно, не существует. Мы в дальнейшем будем пользоваться в основном термином «чувствительность» нервной системы, разумея под этим абсолютную специфическую чувствительность анализаторов (органов чувств).
Следует специально подчеркнуть, что речь идет именно об абсолютной чувствительности, т. е. величине, обратной абсолютному порогу ощущения, а не о различительной (дискриминативной, дифференциальной) чувствительности – величине, обратной порогу различения двух объектов или качеств. Об этом приходится говорить потому, что иногда в дискуссиях о связи между силой нервной системы и чувствительностью два указанных – совершенно различных – содержания последнего термина смешиваются, что ведет к утрате предмета обсуждения, к неточным аргументам и к неправильным заключениям.
Что касается различительной чувствительности, то предпринятые до сих пор попытки связать эту существенную психофизиологическую характеристику со свойствами нервной системы, в частности с предполагаемой «концентрируемостью», способностью нервного процесса к концентрации, не привели к сколько-нибудь ясным результатам (М.Н. Борисова, 1959). Возможно, что это связано с высокой упражняемостью, тренируемостью различительных порогов (Б.М. Теплов, 1947; М.Н. Борисова, 1957), которые, таким образом, вряд ли являются функцией столь устойчивых черт организации нервной системы, как ее основные свойства.
Но вернемся к гипотезе Б.М. Теплова. Она была впервые высказана в печати в 1955 г. В то время эта исключительно плодотворная 'идея действительно оставалась лишь гипотезой, хотя и опиралась на некоторые приводимые разными авторами (И.В. Виноградов, 1933; М.С. Колесников, 1953) наблюдения, указывавшие на повышенную интенсивность и чрезвычайно трудное угашение ориентировочных рефлексов у собак слабого типа нервной системы (возможно, однако, что эти особенности ориентировочного поведения обусловлены не чувствительностью слабого типа, а недостаточной динамичностью тормозного процесса у изученных животных).
Однако за 10 лет, прошедших с этого момента, было накоплено достаточное количество данных, чтобы считать взаимосвязь между абсолютной чувствительностью и силой нервной системы экспе-риментально установленным фактом. Эти данные добыты как в лабо-ратории Б.М. Теплова на людях, так и в некоторых других научных учреждениях на животных. Изложим сначала материалы работ пер-вой группы, а затем остановимся на сообщениях авторов, работавших с животными.
Уже первоначальная проверка гипотезы (В.Д. Небылицын, 1956, 1959а) дала вполне определенные результаты. В ходе этой проверки испытуемые были проведены по трем силовым методикам. Одна из них – индукционная, вариант «кофеин», другая – угашение с подкреплением, фотохимический вариант, а третья еще не была описана на страницах настоящей работы. Ее суть заключается в измерении сдвигов абсолютной чувствительности под влиянием разных доз кофеина (В.Д. Небылицын, 19576). Основанием для ее более детальной разработки послужили некоторые экспериментальные наблюдения над индивидуальными особенностями реакции на кофеин в сфере органов чувств. Литературные данные на этот счет довольно противоречивы, к тому же их не очень много. Укажем в этой связи на работы X. Розе и И. Шмидта (Н. W. Rose, I. Schmidt, 1947), С.И. Субботника (1945), С.А. Брандиса (1938), С.В. Кравкова (1939), К – Трэмэ-ла и др. (К. G. Troemel et al., 1951), исследовавших влияние кофеина на зрительные пороги, а также Р.И. Левиной (1953) и Ю.А. Клаас (1956), изучавших влияние кофеина на слуховые пороги.
Ни в одной из упомянутых работ – кроме разве работы Р.И. Левиной – не делается попытки объяснить или хотя бы как-то связать действие кофеина с особенностями высшей нервной деятельности. Между тем основанием для подобной попытки могло бы служить хотя бы то, что в павловских лабораториях кофеиновая проба при использовании условнорефлекторного метода явилась в конечном счете самым надежным и самым удобным индикатором силы нервной системы.
Техническая сторона описываемой методики очень проста. После установления фонового уровня чувствительности испытуемый получал чистый кофеин в растворе; в первом опыте доза составляла 0,05, во втором – 0,1, в третьем – 0,3 г.
После 20‑минутного перерыва измерение порогов возобновлялось и продолжалось в зависимости от характера изменений чувствительности 30 – 50 мин, с интервалами 2 мин.
Опыты с применением кофеина проводились через день.
Обоснованием валидности этой методики служило сопоставление ее результатов с данными, получаемыми при помощи референтных методик – индукционной и угашения с подкреплением. Первоначально (В.Д. Небылицын, 1956) индикатором силы по этой методике была принята величина сдвигов чувствительности в сторону повышения последней, а именно: отсутствие изменений чувствительности или небольшие ее сдвиги, лежащие в пределах 30 % от фона, были квалифицированы как признак силы нервных клеток, большие же сдвиги чувствительности – до 300 % и более от фона – были истолкованы как проявление слабости нервных клеток.
Однако по мере накопления экспериментальных данных мы должны были ввести сюда некоторые дополнения. Дело в том, что у части испытуемых прием кофеина вызывает не повышение, а понижение чувствительности, достигающее подчас довольно значительных величин, что можно наблюдать как на зрительном, так и на слуховом анализаторе. По результатам сопоставления с данными референтных методик эти испытуемые были признаны «слабыми», в то же время у «сильных» испытуемых случаев снижения чувствительности после приема кофеина не наблюдалось совершенно.
Мы могли заключить отсюда, что показателем слабости нервной системы по этой методике является или сильное повышение чувствительности, или снижение ее (независимо от величины этого снижения). У лиц же с сильной нервной системой прием кофеина или не вызывает никакого изменения чувствительности, или вызывает сравнительно небольшое ее повышение.
Возвратимся теперь к первой экспериментальной работе по определению связи между чувствительностью и силой нервной системы. У всех 37 испытуемых были измерены абсолютные зрительные пороги; данные о слуховой чувствительности были получены от 25 испытуемых. К сожалению, не все испытуемые были проведены по каждой из трех методик определения силы нервной системы. Сопоставление данных о чувствительности и силе шло раздельно для двух анализаторов – зрительного и слухового[31]. У 33 испытуемых сила нервных клеток зрительного анализатора определялась, по крайней мере, двумя экспериментальными способами, а у 11 из них исследование силы было проведено при помощи всех трех методик.
В экспериментальной серии по зрительному анализатору нам встретились лишь отдельные случаи расхождения между результатами испытания силы нервной системы различными методиками. В 91 % всех случаев было получено совпадение результатов, что дало нам право разбить всех испытуемых по суммарной оценке силы нервных клеток в зрительном анализаторе на две основные группы. В одну из групп вошли 15 человек, обнаруживших слабость или тенденцию к слабости нервных клеток, в другую – 22 человека, показавших больший или меньший уровень собственно силы нервной системы. Теперь мы могли статистически сравнить средние показатели абсолютной чувствительности, вычисленные для той и другой группы. Критерий t оказался равен 7,09, р < 0,001, что означало существование весьма четко выраженной зависимости между силой нервной системы и абсолютными порогами.
В опытах на слуховом анализаторе были использованы две методики определения силы нервных клеток: одна из них – изменение чувствительности под влиянием кофеина, другая – угашение с подкреплением, где в качестве условного использовался звуковой раздражитель. У 11 испытуемых сила нервных клеток определялась обоими способами, у 13 она была определена только по влиянию кофеина на чувствительность, у 1‑го испытуемого – только при помощи угашения с подкреплением.
Сопоставление обеих методик приводит нас к выводу, что с их помощью получаются в основном одинаковые результаты. Это, во всяком случае, справедливо для 10 испытуемых из 11.
По суммарной оценке силы нервных клеток все 25 испытуемых могут быть распределены по двум группам. Средняя чувствительность группы «сильных» составляет 96 усл. ед., «слабых» – 162 усл. ед. Вычисление критерия t дает его величину, равную 3,56 (р < 0,01).
Таким образом, разность средних значений чувствительности для слухового анализатора статистически менее значима, чем для зрительного, хотя и вполне удовлетворяет критериям обоснованного вывода. Меньшая величина t здесь обязана своим происхождением кроме меньшего числа испытуемых еще и двум случаям явных исключений из общей картины, заключающихся в сочетании высокой чувствительности с высокой же силой нервных клеток. Причиной этих исключений, возможно, является несовершенство той единственной силовой методики, при помощи которой данные испытуемые были исследованы (непосредственное влияние кофеина на чувствительность).
Но независимо от этого нужно заметить и подчеркнуть, что отрицательная связь между чувствительностью и силой, как это следует из экспериментальных материалов, отнюдь не является функциональной (в математическом смысле этого термина). В материалах не так уже редко встречаются случаи сочетания высокой чувствительности с высокой силой нервной системы и наоборот – низкой чувствительности с явной слабостью нервных клеток. Очевидно, дело здесь не сводится к несовершенству методик, хотя, вероятно, и этот фактор может играть определенную роль. Скорее, можно думать о влиянии факторов функционального состояния организма, надо полагать, способных значительно видоизменять характер функции, особенно такой тонкой, как абсолютная чувствительность.
Имеется целый ряд экспериментальных свидетельств в пользу того, что колебания порогов происходят непрерывно в течение как самых малых, так и весьма длительных промежутков времени (П.П. Лазарев, 1923; К. X. Кекчеев, 1946; П.Г. Снякин, 1951). Чувствительность не остается постоянной даже в отрезки времени, измеряемые секундами и минутами; она изменяется от одного опытного дня к другому. Можно предполагать также существование более медленных, но, возможно, более крупных сдвигов в периоды времени, измеряемые месяцами или даже годами. Это же, вероятно, относится и к функции выносливости, работоспособности нервной ткани, составляющей главное содержание силы нервной системы.
Все это приводит к тому, что измерение тесноты связи между двумя переменными, из которых одна представляет собой абсолютную чувствительность, а другая – выраженные в количественной форме результаты определения силы нервной системы, никогда не дает очень высоких значений. Коэффициенты корреляции в лучшем случае достигают лишь величины порядка 0,7; это, конечно, совсем не мало, но если учесть, что общий для двух переменных фактор даже в этом случае «отвечает» примерно лишь за 50 % дисперсии, станем ясно, сколь велика доля общей дисперсии, обусловленная влиянием неучтенных причин.
Итак, первоначальная проверка гипотезы Б.М. Теплова о связи между чувствительностью и силой подтвердила эту гипотезу и вместе с тем показала, что зависимость между двумя этими параметрами нервной системы носит статистический характер, выступая отнюдь не в виде функционального соотношения, а в виде корреляции. В работах, выполненных позже с использованием корреляционной и факторной техники анализа получаемых соотношений, были неоднократно подтверждены как существование самой зависимости, так и ее статистический характер.
В совместной работе В. И, Рождественской и др. (1960), посвященной сопоставлению всех принятых к тому времени в лабораторч:* индикаторов силы нервной системы, в сопоставление были включены и показатели абсолютных порогов – зрительных и слуховых. Особенностью этой работы явилось то, что 38 испытуемых из 40 были проведены по всем методикам и, таким образом, можно было подсчитать коэффициенты корреляции каждого показателя с каждым другим. В данный момент нас интересуют корреляции между показателями чувствительности и референтными индикаторами силы нервной системы. Последних, если к ним отнести и влияние кофеина на чувствительность, было 10. Перечислим их:
1. Угашение с подкреплением фотохимического условного рефлекса, при зрительных условных раздражителях, без кофеина.
2. Угашение с подкреплением фотохимического условного рефлекса, п'ри зрительных условных раздражителях, с применением кофеина.
3. Индукционная методика, вариант «форма кривой».
4. Индукционная методика, вариант «утомление».
5. Индукционная методика, вариант «повторение».
6. Индукционная методика, вариант «кофеин».
7. Измерение действия кофеина на зрительную чувствительность.
8. Угашение с подкреплением фотохимического условного рефлекса, при слуховых условных раздражителях, без кофеина.
9. То же, с кофеином.
10. Измерение действия кофеина на слуховую чувствительность.
Как видно из этого перечисления, 7 индикаторов относятся к зрительному анализатору (1 – 7), а остальные три – к слуховому (8 – 10). Коэффициенты корреляции рангов между референтными индикаторами силы и показателями порогов приводятся в табл. 24, в которой номера методик соответствуют только что данному перечислению.
При рассмотрении таблицы обращает на себя внимание один примечательный момент: коэффициенты корреляции для показателей зрительных порогов почти во всех случаях выше, чем для слуховых, а если ниже, то на весьма незначительную величину; среди первых нет ни одного незначимого коэффициента, среди вторых – их четыре. С этим отличием зрительного анализатора от слухового мы, собственно, уже сталкивались выше, когда отмечали, что статистическая значимость разности между средними значениями чувствительности «сильных» и «слабых» для зрительного анализатора оказывается выше, чем для слухового. Теперь, при корреляционном анализе материала, это явление повторяется. В чем его причины?
В качестве предположительного объяснения некоторых из этих причин можно указать на морфофизиологические различия в организации периферических сенсорных аппаратов зрения и слуха. Если первичные процессы зрительного анализатора имеют с самого начала фотохимическую, а затем нейроэлектрическую природу, то в слуховом анализаторе первая фаза обработки энергии стимула, вплоть до раздражения так называемых волосковых клеток, приходится на чисто механические процессы. Эта разница, в общем, видимо, соответствует различиям в качестве самих физических энергий, служащих адекватными раздражителями для зрительного и слухового рецепторов. Можно предполагать, что механические свойства периферии слухового анализатора более подвержены влиянию различных условий существования организма, чем химические и нейрофизиологические свойства сетчатки, тем более что сетчатка – это закрытый орган, надежно защищенный глазным яблоком, а улитка и особенно органы среднего уха расположены в непосредственной близости к наружному слуховому проходу. Это, возможно, и обусловливает большую частоту несоблюдения правила связи между чувствительностью и силой нервных клеток в слуховом анализаторе, приводящую к тому, что корреляции между слуховой чувствительностью и силовыми методиками для слухового анализатора, как правило, ниже, чем между зрительной чувствительностью и результатами определения силы в зрительном анализаторе.
|
|
|
|
|
Дата добавления: 2015-06-26; Просмотров: 432; Нарушение авторских прав?; Мы поможем в написании вашей работы!