КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Белоктарды топтастыру. 2 страница
Сондықтан да, барлық жасушаларда немеее тек эмбрионалдық жасушаларда (ствол жасушаларында), ДНҚ-ның толық репликацияланбаған учаскелері қалпына келуі қажет. Бұл қызметті теломеразалар атқарады. Олар қалай әрекет етеді.
Теломеразалар әрекетінің тетіктері
Теломеразалар әрбір теломералардың G-тізбегін ұзартады. Теломеразалармен 450 нуклеотидтерден тұратын теломеразалық РНҚ байланысқан. Оның ортаңғы қысқа учаскесі 1,5 теломерлік қайталануға комплиментарлы болады (26 сурет).
Осы РНҚ-ның сол жағындағы триплет (АУЦ) ДНҚ-ның G -тізбегенің шеткі теломерлік жартықайталанумен байланысу (гибридтену) үшін пайдаланылады. Қалған гексонуклеотид (ЦЦААУЦ) G -тізбекті 31 ұшынан ұзарту үшін матрица ретінде қызмет атқарады.
Теломеразалар қызметі, өте қызық-таңқаларлық, ол қысқа, жаңадан синтезделген тізбекті ұзартпай, ескі аналық (матрицалық) ұзын тізбекті ұзартады (6-сурет)
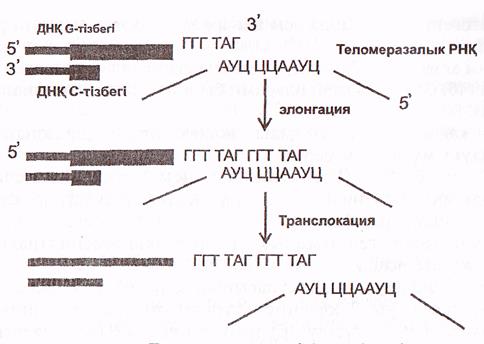
6-сурет. Теломераза әрекетінің тетіктері (Мушкамбаров, Кузнецовтан, 2003)
Аналық (ескі, ұзын) тізбектің 31 ұшына теломераза бірізділікпен бірнеше ондаған, тіпті жүздеген, гексонуклеотидтерді (ГГТТАГ) жалғайды (элонгация, транслокация). Осыдан кейін біршама ұзарған (аналық) тізбек тағы бір Оказаки фрагментінің синтезделуі үшін матрица қызметін атқарады.
Ол жоғарыда сипатталған ДНҚ синтезі сияқты жүзеге асады. Алғаш аналық тізбектің 31 ұшында праймаза «РНҚ ұйытқының синтездейді, сосын ДНҚ-полимераза β теломерлік қайталануларға комплиментарлы дезоксинуклеотидтерді ұйытқыға жалғайды. Фрагменттің өсуі 5'—>3' бағытында жүреді, ал оның аяқталуы алдыңғы фрагменттің 51 ұшымен түйіскенде ғана жүзеге асады. Синтезделген фрагменттің ДНҚ тізбегіне жалғануын ДНҚ-лигаза қамтамасыз етеді. Экзонуклеаза жаңа тізбектегі «РНҚ-ұйытқыны» алып тастайды. Нәтижеде ДНҚ қос тізбегі бұрынғы ұзындығына ие болады.
№7 дәріс
Тақырыбы: ДНҚ транскрипциясы немесе РНҚ синтезі
Жоспар: 1. Жалпы мәліметтер
2. Транскрипцияның жалпы сипаттамасы
3.Транскрипция факторлары
ДНҚ молекуласында генетикалық ақпарат болатыны белгілі, ол:
-ағзаның барлық акуыздары және РНҚ молекулалары туралы ақпарат;
-онтогенез барысында осы ақпараттың жүзеге асуының реті туралы ақпарат.
Адам ағзасының барлық дене жасушаларында хромосома жиынтығы бірдей (46) болғандықтан олардың бәрінде бірдей генетикалық ақпарат болады.
Бұл жағдай, яғни генетикалық эквиваленттілік, диплоидты ағзаларды клондауға мүмкіндік береді.
Өзімізге белгілі ДНҚ репликациясы нәтижесінде генетикалық ақпарат екі еселенеді және олар жаңадан түзілген жасушаларға тепе-тең мөлшерде беріледі.
Бұдан басқа, генетикалық ақпарат экспрессияланады яғни әрі қарай жүзеге асады.
Белгілі бір ақуыз молекуласының құрылымы туралы ақпараттың экспрессиялануы 2 кезеңнен тұрады: а) транкрипция —жасуша ядросында а-РНҚ түзілуі; б) трансляция —аРНК, ақпараты негізінде рибосомаларда ақуыз молекуласының синтезделуі.
Транскрипция дегеніміз — ДНҚ молекуласындағы генетикалық ақпараттың РНҚ молекуласына көшіріліп жазылуы, яғни РНҚ синтезделуі болып табылады.
Егер ДНҚ репликациясы жасушаның бөлінуіне байланысты болатын болса, яғни тек бөлінуші жасушаларда байқалатын болса, транскрипция үдерісі барлық ядролы жасушаларда, бөлінетін және белінбейтін, байқала береді.
Бөлінетін жасушаларда ол митоздық циклдың кез-келген уақытында жүреді және ДНҚ молекуласының бір учаскесінің транскрипциялануы көп рет қайталанып жүруі мүмкін.
Транскрипция немесе РНҚ синтезі үшін құрылыс материалдары болып рибонуклеозидтрифосфаттар (рАТФ, рГТФ, рЦТФ, рУТФ) саналады. РНҚ тізбегіне қосылған кезде олар 2 фосфат қалдығын пирофосфат күйінде бөліп шығарып, босаған энергияны нуклеотидтер арасындағы фосфодиэфирлік байланыстың түзілуіне жұмсайды.
Еркін рНТФ —> рРНҚ тізбегіндегі қалдығы +пирофоефат
РНҚ синтезі промоторлық учаскеден басталып терминаторлық учаскемен аяқталады. РНҚ синтезі жүруі үшін ДНҚ молекуласының кем дегенде 2 өрімінде ДНҚ жіпшелері бір-бірінен ажырасулары қажет, ширатылған күйінде транскрипция жүрмейді.
РНҚ синтезі ассиметриялық құбылыс, яғни бір-бірінен ажырасқан ДНҚ жіпшелерінің тек біреуі ғана РНҚ синтезі үшін матрица (қалып) қызметін атқара алады. РНҚ синтезі консервативтік құбылыс, яғни транскрипция аяқталғаннан кейін ДНҚ молекуласы ширатылып, бастапқы күйіне келеді.
РНҚ синтезін РНҚ-полимераза — ферменті өз бетінше жүргізеді. Яғни ешқандай ұйытқының қажеті жоқ.
Транскрипция өнімі болып жетілмеген РНҚ-лар: пре-аРНҚ, пре-тРНҚ, пре-рРНҚ-лар саналады. Олар ядрода пісіп жетіледі (процессинг).
Гендердің экспрессиялануына көптеген ақуыздар әсер етеді. Соңғы жылдары транскрипциялық фактор қасиетіне ие көптеген ақуыздар анықталды. Олар бір-бірімен, не ДНҚ молекуласының реттеуші учаскелерімен (промотор, энхансерлер), не басқа да заттармен (лиганда) әрекеттесіп гендердің белсенділігіне әсер етеді.
№8 дәріс
Тақырыбы: ДНҚ молекуласының реттеуші
учаскелерімен байланысатын ақуыздар
ДНҚ-мен байланысатын ақуыздар саны өте көп. Құрылымдарына қарай оларды бірнеше топқа топтастырады:
а) «Ширатпа-бұрылым-ширатпа» мотивтері болатын ақуыздар.
Бұл ақуыздардың ДНҚ-ны танитын, онымен байланысатын учаскелері ілмек арқылы қосылған екі α- ширатпадан тұрады. Олардың біреуі белгілі бір нуклеотидтер бірізділігімен спецификалық әрекеттесіп, ДНҚ молекуласымен тікелей байланысады
(7-сурет).
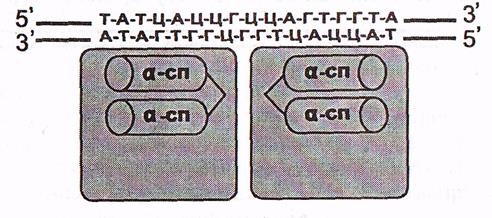
Ақуыздың екі субъединицасы
7-сурет. «Ширатпа-бұрылым-ширатпа» мотиві болатын ақуыздың ДНҚ-мен әрекеттесуі (Мушкамбаров, Кузнецовтан, 2003)
б) Гомеодомендері болатын ақуыздар.
бұларға эукариоттардың эмбриональдық дамуына жауапты гомейотикалық гендердің өнімдері жатады. Осы ақуыздар арқылы кейбір гендерді іске қосып, кейбіреулерін істен шығарып ағзалардың дамуы реттелінеді (8-сурет).
ГОМЕОДОМЕНДЕР
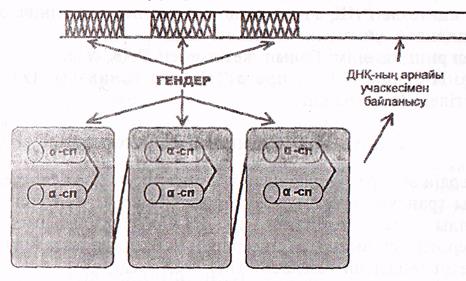
8-сурет. Гомеодомендік ақуыздар (Мушкамбаров, Кузнецовтан, 2003)
Бұл ақуыздардың ерекшелігі-гомеодомендер деп аталатын біртипті домендердің болуы.
Гомеодомен, шамамен, 60-тай аминқышқылдарынан құрылған, құрылысы жағынан прокариоттардың ақуыз-репрессорларына ққсас
болады.
в) «Мырыш саусақтары» болатын ақуыздар.
Бұл ақуыздардың саусақтерізді құрылымдары болады. Олар мырыш атомының 2 цистеин және 2 гистидин аминқышқылдарының қалдықтарымен 4 байланыс пайда ету арқылы тұрақтанады (9-сурет).
ДНҚ
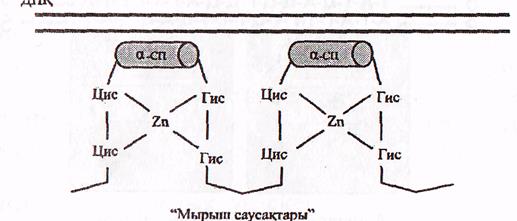
"Мырыш саусақтары"
«Мырыш саусақтары бар» ақуыздардың ДНҚ мен әрекеттесуі (Мушкамбаров, Кузнецовтан, 2003)
Саусақтың сыртқы бетінде ДНҚ-ның белгілі бір нуклеотид бірізділігін арнайы танитын α-ширатпа орналасқан. Саусақтардың саны әртүрлі ақуыздарда түрліше болады.
Бұл топқа эукариоттардың көптеген реттеуші ақуыздары, атап айтқанда стероидтық гормондардың жасушаішілік ақуыз-рецепторлары жатады.
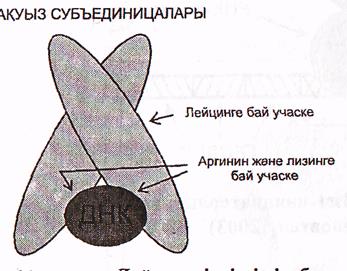
г) Лейцин «ілдіргіші» бар ақуыздар.
Бұл топқа бір-бірімен Лейцин қалдықтары арасындағы гидрофобтық байланыстар арқылы қосылған 2 субъединицадан (бөлшектен) тұратын ақуыздар кіреді.
ДНҚ-мен байланысатын учаскеде негіздер қасиетіне ие аминқышқылдар-аргинин және лизин көптеп кездеседі (30-сурет).
Бұл топқа эукариоттардың көптеген транскрипциялық факторлары кіреді.
№9 дәріс
Тақырыбы: Транскрипцияның жалпы факторлары
Жоспар: 1. р-53 ақуызы-транскрипция факторы
2. Транскрипция тетіктері (механизмдері)
Транскрипцияның жалпы факторлары РНҚ полимераза ферментінің промотормен байланысуын қамтамасыз ететін ақуыздар.
10-сурет. Леицин «ілдіргіші» бар ақуыздың ДНҚ мен әрекеттееуі
(Мушкамбаров, Кузнецовтан, 2003).
Бұл ақуыздардың өздеріде промотормен байланысады.
Промоторлық учаскеде бокстар деп аталатын ерекше нуклеотидтер бірізділігі кездеседі, мысалы эукариоттарда ТАТА бокс жиі кездеседі, сонымен бірге сиректеу ГЦ-, ЦТ-бокстары да белгілі. Әртүрлі дромоторларда олардың орналасу реті түрліше; кейбір жағдайларда ГЦ-бокс, ТАТА-боксқа дейін орналасса, екінші біреулерінде керісінше.
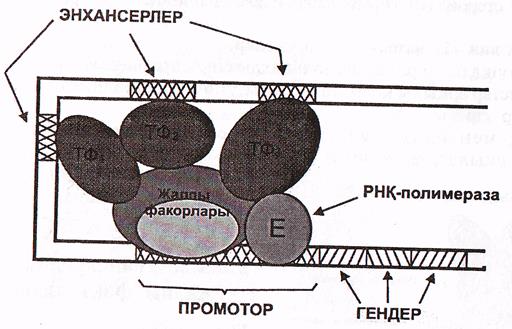
Промотордың ТАТА-боксымен алғаш ТВР-ақуыз байланысады. Бұл бірнеше ТАҒ-ақуыздардың жалғануын инициациялайды.
Бұл ақуыздар (ТВР және ТАҒ ақуыздары) - транскрипцияның жалпы факторлары деп аталады, себебі олар барлық жасушаларда кездеседі және көптеген гендердің экспрессиялануы үшін міндетті түрде қажет. Осы ақуыздар кешенін ТҒ ІІ Д деп те атайды.
Осы ақуыздар РНҚ-полимераза ІІ-нің промотормен байланысуы
үшін қажет. Промотормен байланысқан ақуыздар кешенін негізгі инициаторлық кешен деп атайды. ТҒІІД дан басқа ТҒІІА, ТҒІІВ, ТҒІІЕ, ТҒІІН деп аталатын транскрипсцияның жалпы факторлары да белгілі. Олар әртүрлі промоторлармен байланысады (11-сурет).
Транскрипцияның негізгі инициаторлық кешені (Мушкамбаров, Кузнецсовтан, 2003)
р-53 ақуызы жасушаның көптеген құбылыстарын реттеуге қатынасады және оның қызметі сан алуан:
1. Жасуша құрылымдарының бұзылыстарына жауап ретінде р-53,
не оның гені активтенеді.
2. р-53 ақуызы 3 топ гендердің белсенділігін реттейді:
а) Р-21, GAAD 45 т.б. жасуша бөлінуін тоқтататын гендерді
активтеңдіреді;
б) апоптозды іске қосатын ВАХ, КіІІег/ DR5, РІG т.б. гендерді
активтендіреді;
в) апоптозды тежейтін BCL2, RELA гендерін репрессиялайды;
г) ангиогенезді тежейтін TSP1, ВАІІ т.б. гендерді активтендіреді
яғни ақуыз р-53 көмегімен жасуша өз құрылымдарының бұзылыстарына төмендегідей жауап қайтарады:
-митоздық циклдың белгілі бір сатысында оның әрі қарай бөлінуін тоқтатып бұзылыстарды жөндеуге мүмкіндік жасайды; -немесе әрі қарай бөлінуін мүлдем тоқтатып жасуша қартаю кезеңіне өтеді;
-немесе апоптоз іске қосылады да жасуша өліп жойылады.
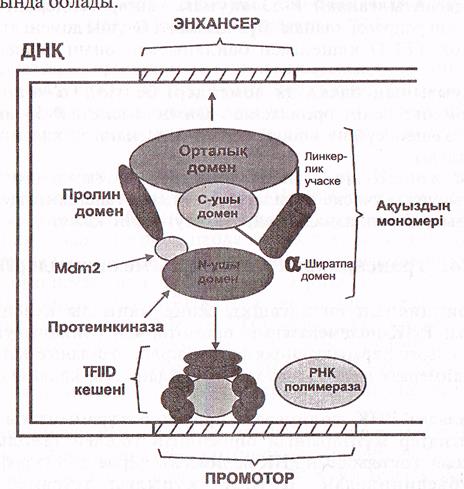
Р-53 молекуласы 392 аминқышқылдарынан құрылған және көлемі, қызметтері әртүрлі 6 доменнен тұрады (12-сурет).
Орталық домен ең үлкен домен (200 аминқышқылы кездеседі) ген нысананың энхансерлерін танып білу қызметін атқарады және олармен байланысады.
Ақуыздың N ұшында N ұшы домені транскрипцияның жалпы факторларымен, яғни ТҒІІD кешенімен әрекеттеседі.
Бұл байланысулар басқа да көптеген факторлардың (домендердің) бақылауында болады.
12-сурет. Р-53 ақуызының құрылысы (Мушкамбаров, Кузнецовтан, 2003)
N доменде ақуыз—ингибитор Мdm -2-мен байланысатын локус болады. Ол ТҒIID кешенін бастырмалайды. Осы жерде серинтрионин аминқышқылдары қалдықтары көптеп кездеседі және олар протеинкиназалармен фосфорланады. Бұл киназалар ДНҚ молекуласының құрылысы бұзылған кезде активтенеді де, Мdm 2-ингибиторына әсер етіп, оны пассив күйіне көшіреді, нәтижеде 14-домен ТҒIID кешенімен әрекеттесу қабілетіне ие болады.
Орталық доменнің энхансерлермен әрекеттесуі де бақылауда болады, ол модификациялану арқылы жүзеге асады, бірақ бұл кезде орталық домен емес С-ұшы-домені модификацияланады және ол көптүрлі болып келеді: фосфорлану, ацетильдену, гликозилдену т.б.
Егер С-ұшы-домені модификацияланбаса, орталық домен ДНҚ-нысанамен (энхансерлер) әрекеттесе алмайды.
Жоғарыда айтылғандай Р-53 ақуызы кейбір гендердің—ВCL2, RELA, әрекетін репрессиялайды. Бұл қызметті С-ұшы домені атқарады. Бұл кезде ол ТҒIID кешенімен байланыеып, оның белсенділігін тежейді.
Р-53 ақуызының басқа да домендері белгілі: α-ширатпалы домен С-доменге дейін орналасқан. Оның қызметі-Р-53 ақуызын тетрамерлік кешен күйіне көшіріп оның белсенділігін қалыптастыру болып табылады.
Орталық жөне α-ширатпалы домендер арасында линкерлік (байланыстырушы) учаске орналасқан. Ол енді ғана синтезделген Р-53 ақуызының сцитоплазмадан ядроға өтуі үшін қажет.
Транскрипцияның ең алғашқы және маңызды кезеңі-оның инициациясы: РНҚ-полимеразаның промотормен байланысуы және алғашқы нуклеотидтераралық (фосфодиэфирлік) байланыстың түзілуі.
РНҚ полимераза мен промотордың байланысуы қалайша жүзеге асады?
Бактерияларда РНҚ полимераза промотор құрамындағы белгілі бір нуклеотидтер жұптарының бірізділігін тікелей таниды, мыс: Прибнов боксын. Бактерияның РНҚ полимераза ферментінің коферменті 3 түрлі субъединицадан - αββ1, қүрылған тетрамер болып табылады. Ол өздігінен промотормен байланыса алмайды, ал егер оған ерекше ақуыз- σ -фактор жалғанса онда σ -фактордың қатынасуымен РНҚ-полимераза ферменті промотордың Прибнов боксын танып, онымен байланысады да транскрипцияны бастайды (13 сурет).
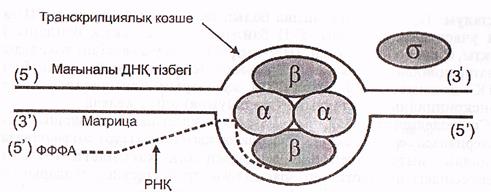
13-сурет. Бактерия ДНҚ-сының транскрипциялану жобасы (Мушкамбаров, Кузнецовтан, 2003)
Эукариоттарда промотормен көптеген ақуыздар байланысады, мыс: ТҒІІD, ТҒІІА, ТҒІІС, ТҒІІЕ, ТҒІІН кешендері, сонымен қатар ген транскрипңиясының инициасиялауы үшін осы геннің энхансерлерімен байланысатын басқа да транскрипция факторлары (мыс: ақуыз Р-53) қажет.
РНҚ полимераза промотормен байланысып, ДНҚ молекуласының локальды денатурациялануын, яғни ДНҚ тізбектерінің 1,5-2 ширатпа ұзындығында бір-бірінен ажырауын туғызады. Осылайша транскрипциялық «көзше» пайда болады және «көзшедегі» ДНҚ-ның матрицалық тізбегінде орналасқан нуклеотидтердің р →НМФ-тармен комплиментарлы жұптасуына мүмкіндік туады.
Жаңадан синтезделуші РНҚ тізбегіне алғашқылардың бірі болып пуриндік нуклеотид-АТФ не ГТФ қосылады жөне ол өзінің 3 фосфат қалдығын сақтап қалады. Содан кейін екінші нуклеотидпен алғашқы 5'-3-фосфаттық байланыс түзіледі. Осыдан кейін бактерияларда б-фактор РНҚ-полимеразамен байланысын үзіп, түсіп қалады.
Инициациядан кейінгі кезең - элонгация: синтезделуші пре-РНҚ тізбегінің жайлап ұзаруы терминациялық учаскеге дейін жалғасады. РНҚ синтезінде 1 секундта шамамен 30 нуклеотид жалғанады.
Жалпы алғанда транскрипция қателіксіз жүреді, себебі ол матрицалық (қалып), комплиментарлық принциптерге негізделінеді, бірақ кейде 2х104 нуклеотидтен біреуі қате жұптасуы мүмкін. Бұл қателіктер мутацияға алып келеді, сондықтан олар дер кезінде эндонуклеазалар арқылы жөнделіп отырады.
Транскрипцияның соңғы кезеңі терминация, немесе транскрипцияның аяқталуы. Терминацияға сигнал болып геннің аяқ жағындағы ГЦ-ға бай учаскелері саналады. Г-Ц байланысы (3 сутектік байланыс) мықты, берік болғандықтан ДНҚ-ның осындай учаскесінің локальды денатурациялануы (екі жіпшесінің ажырасуы) қиындай түседі. Бұл РНҚ—полимераза ферментінің жылжуын баяулатады және транскрипцияның тоқталуына (аяқталуына) алып келеді. Синтезделген пре-РНҚ-ның ДНҚ молекуласынан босанып шығуын бактерияларда ерекше ақуыз- Rһо-фактор қамтамаеыз етее, эукриоттарда жаңадан еинтезделген пре-РНҚ-ның аяқ жағындағы ГЦ-бай учаскесіндегі нуклеотидтер арасындағы әрекеттесулер салдарынан пайда болатын «ілмекшелер» («шпильки») атқарады.
№10 дәріс
Тақырыбы: Транскрипцияның алғашқы өнімдері
Жоспар: 1. Транскрипцияның алғашқы өнімдері
2.Пре-РНҚ-лардың пісіп жетілуі— процессинг
Транскрипция нәтижесінде эукариоттарда жетілмеген пре-РНҚ (пре-аРНҚ, пре-тРНҚ, пре-рРНҚ) синтезделінеді, себебі эукариоттар генінің құрылысы бактерияларға қарағанда күрделірек, яғни ол экзон-интрондық құрылысқа ие болады және транскрипңия кезінде пре-РНҚ-ларда экзондық-интрондық учаскелері түгел көшіріліп жазылады (34-сурет).
а) пре-аРНҚ-жетілген а-РНҚ-ларға қарағанда әлде қайда ұзын
болады, себебі олардың құрамына спейсерлер (реттеуші, құрылымдық
қызметтер атқаратын ДНҚ учаскелері), мағыналы ДНҚ учаскелері-
экзондар, мағынасыз учаскелері-интрондар кіреді. Сондықтан да пре-
РНҚ-ларды кейде гетерогендік ядролық РНҚ (гя-РНҚ) деп те
атайды.
пре а-РНҚ-лардың келесі ерекшелігі оның 5' ұшында «қалпақшаның» (КЭП), 31 ұшында-поли (А)-фрагменттің болмауы.
б) рРНҚ-ның кластерлі 3 гені біртұтас транскрипцияланады
және синтезделген пре-рРНҚ немесе 45S РНҚ-құрамында жетілген
үш түрлі рРНҚ-ға 18S, 5.8S, 28S РНҚ-сәйкес келетін бірізділіктер
болады. Бұл бірізділіктер спейсерлермен бөлінген, бірақ онда интрондар
болмайды. Сонымен қатар, бұл жерде жетілген рРНҚ-ларда кездесетін
модификацияланған нуклеотидтерде болмайды.
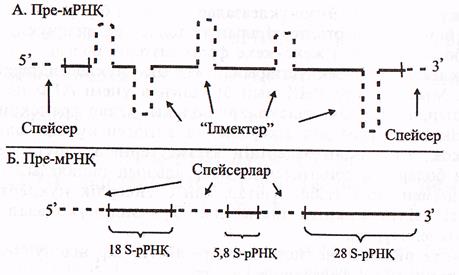
14-сурет Пре-а РНҚ және пре-р РНҚ (Мушкамбаров, Кузнецовтан, 2003)
в) Барлық пре-РНҚ-лардан ерекше пре-тРНҚ—лар тек жетілген бірізділіктерді қамтиды. пре-тРНҚ молекуласының сырт пішіні «жөке ағашының жапырағына» («кленовый лист») ұқсас үшқұлақты болады, бірақ оның жетілген тРНҚ молекуласынан төмендегідей ерекшеліктері белгілі (15-сурет).
-молекуланың екі ұшында және ортасында қосымша бірізділіктер болады;
-минорлық (модификацияланған) нуклеотидтер болмайды;
-акцепторлық ілмекше (ЦЦА) толық қалыптаспаған;
-антикодон өз орнында емес, басқа жерде орналасқан.
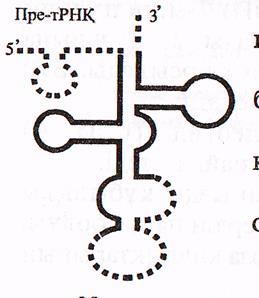
пре-РНҚ молекуласының пісіп жетілуі (процессинг) 3 кезеңге бөлінеді:
- кейбір нуклеотидтердің алынып тасталуы;
- кейбір нуклеотидтердің жалғануы;
- олардың модификациялануы;
Артық нуклеотидтердің алынып тасталуы ерекше нуклеазалар арқылы жүзеге асады. Экзонуклеазалар тізбектің бір ұшынан (31 не 51) бір — бірлеп нуклеотидтерді алып тастайды, ал эндонуклеазалар тізбекті бөлшектеп оны жеке-жеке фрагменттерге беледі.
а) Нуклеазалар тізбек ұштарынан «артық» нуклеотидтерді үзіп
отырады. Мысалы, пре-РНҚ-ның 51 ұшында үнемі АТФ не ГТФ,
ал 31 ұшында ГЦ—бай учаскелері болады. Олар транскрипция
үдерістерінде маңызды рөл атқарады, ал жетілген күйінде олардың
қажеті жоқ, тіпті олар өздерінің қызметтерін атқаруға кедергі
келтірген болар еді, сондықтан да олар алынып тасталады.
б) сонымен бірге тізбек ұштарынан спейсерлік нуклеотидтер
бірізділігі үзіліп алынып таеталады. Бұл-эндонуклеазалардың
қатынасуымен жүреді.
в) 45S-пре-рРНҚ және гистондық пре-аРНҚ-лар эндонуклеазалар
арқылы жеке РНҚ тізбектеріне кесіледі.
г) пре-тРНҚ және барлық пре-аРНҚ-лардың интрондық
бірізділіктері кесіліп алынып тасталады. Сол сияқты, экзондық
бірізділіктер түтас бір тізбекке жалғанады, оны сплайсинг деп
атайды. Мұның іске асырылуы үшін тек қана эндонуклеазалар
емес, лигазалар да қажет.
Сонымен, пре-РНҚ молекуласының пісіп жетілуі (процессинг) барысында оларға көптеген нуклеотидтер транскрипциясыз байланысады (жалғанады).
пре-аРНҚ-ның 51 ұшына «қалпақшаның» (КЭП) 7-метилгуанин және басқа да 3-4 нуклеотидтері қосылып жалғанады, ал 31 ұшына 200-дей нуклеотидтерден тұратын поли (А)-фрагмент қосылады. Бұл үдерісті полиаденилатполимераза ферменті катализдейді.
пре-тРНҚ молекуласының 31 ұшына 3 нуклеотид (Ц, Ц, А) бірінен кейін бірі жалғанып, акцепторлық учаске пайда етеді.
пре-РНҚ-ның пісіп жетілуінің (процессинг) маңызды құбылысы олардың құрамында модификацияланатын нуклеотидтердің пайда болуы. пре-а-РНҚ-да «қалпақша» нуклеотидтерінің рибоза қалдықтарының метилденуі байқалады.
Пре-т-РНҚ-да модификациялану көптүрлі болып келеді, мысалы: уридин қалдығы тотықсызданады (дигидроуридин пайда болғанға дейін), басқалары-изомерленеді (псевдоуридин), үшінші біреулері -метилденеді (метилуридин). Аденозиннің кейбір қалдықтары дезаминденіп инозинге айналады, соңғыларының (инозин) кейбіреулері тағы да метилденеді (метилинозин).
Жоғарыда сипатталған құбылыстар ядрода бірнеше пісіп жетілген РНҚ молекулаларының пайда болуына алып келеді, мысалы:
а) 4 түрлі рРНҚ-ның: 28S рРНҚ; 18S рРНҚ; 5.8S рРНҚ; 5S
рРНҚ;
б) бірнеше ондаған т-РНҚ-ның (әрбір 20 аминқышқылына 1-4 ке
дейін);
в) мыңдаған аРНҚ молекулаларының түзілуіне.
№11 дәріс
Тақырыбы: а-РНҚ-лардың ыдырауы
Жоспар: 1.Бактериялардың а-РНҚ-ның 51 ұшынан ыдырауы
2. Эукариоттар а-РНҚ-ның 31 ұшынан ыдырауы
Бактериялар а-РНҚ-сының ыдырауы 51 ұшынан басталып а-РНҚ синтезделу (5'-3') бағытында жалғасады. Сонымен қатар, бактерияларда ядро болмайды, ал а-РНҚ-лары әжептеуір ұзын, полицистронды болып келеді.
Осылардың бәрі, а-РНҚ-ның бір тізбегінің бір мезгілде 3 түрлі үдерістерге қатынасуына мүмкіндік береді:
- ДНҚ молекуласынан өзінің (а-РНҚ) пайда болуына (транскрипция);
- рибосомаларда трансляциялануына, яғни қызмет етуіне;
- өздігінен ыдырауына, яғни 51 РНҚ-азалар әсерлерінен 51
ұшының біртіндеп қысқаруына (16-сурет).
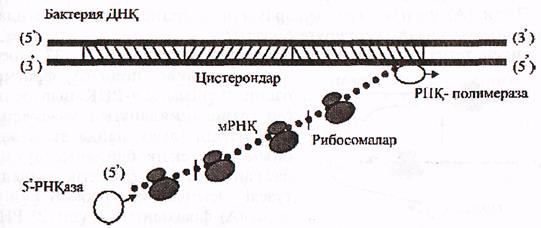
16-сурет. Бактериялар а-РНҚ-ның транскрипциялану, трансляциялану
және ыдырау үдерістерінің қабаттасып келу жобасы.
(Мушкамбаров, Кузнецовтан, 2003)
Эукариоттар а-РНҚ-ның тіршілік ұзақтығы 10 минуттан (қысқа тіршілік ететін а-РНҚ) 2 тәулікке дейін созылады. Қысқа уақыт тіршілік ететін а-РНҚ-ларға реттеуші ақуыздар а-РНҚ-лары жатады. Эукариоттар а-РНҚ-сы моноцистронды болып келеді. Олардың ыдырауы бактериялардағыдай 51 РНҚ азалар көмегімен емес, 3' РНҚ-азалар арқылы жүзеге асады, яғни әдетте ол 3^-ұшындағы 200 нуклеотидтерден құрылған поли-(А)-фрагменттен басталады.
Демек, бұл фрагмент ДНҚ теломералары сияқты буферлік қызмет атқарады. Оның ұзындығының жайлап қысқаруы белгілі бір уақытқа дейін нуклеин қышқылының маңызды, кодтаушы бөлімдерін ыдыраудан сақтайды. а-РНҚ-ның поли- (А)-фрагменті 31 РНҚ-аза ферменті әсерлерінен үнемі қысқармайды, оның қызметтік белсенділігіне сай, мезгіл-мезгіл қысқарады. а-РНҚ-ның әрбір трансляциясы аяқталғаннан кейін рибосома поли-(А)-фрагменттен 10-15 нуклеотидтерді үзіп алып тастайды. Осы фрагментте 50-ге жуық нуклеотидтер қалған кезде а-РНҚ РНҚ-аза әсеріне ілігіп тез ұшырайды.
|
|
Дата добавления: 2015-06-04; Просмотров: 2152; Нарушение авторских прав?; Мы поможем в написании вашей работы!