КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Внутри- и межвидовые отношения в биогеоценозе
|
|
|
|
О чем свидетельствует сопряженность?
Если речь идет о положительной сопряженности, то она может иметь место в двух случаях:
- виды «приспосабливаются» друг к другу настолько, что чаще встречаются друг с другом (свиты видов тех или иных типов леса, в с/х чеснок и морковь), чем порознь
- оба вида сходны по своим экологическим особенностям и чаще обитают вместе потому, что в пределах одного фитоценоза условия оказываются более благоприятными для обоих видов (виды одних и тех же ярусов).
При отрицательной сопряженности она может зависеть от того, что в результате межвидовой борьбы:
- оба вида стали антагонистами (не надо рядом сажать землянику и морковь; волжанка, вейник – угнетают своих соседей по эконише);
- у видов различно отношение к влаге, освещению и другим факторам среды в пределах фитоценоза (растения разных ярусов и разные парцелл).
а) Дифференциация ценопуляций
Лесоводам давно известно, что число стволов деревьев на единицу площади с возрастом насаждений уменьшается. Чем более светолюбива порода и чем лучше условия произрастания, тем быстрее самоизреживается древостой. Гибель деревьев особенно интенсивна в первые десятилетия и постепенно уменьшается с увеличением возраста леса. Это наглядно показано в таблице 2.
Таблица 2
Уменьшение общего числа стволов с возрастом (по Г. Ф. Морозову, 1930)
| Возраст в годах | Число стволов на 1 га | ||
| Буковый лес на раковистом известняке | Буковый лес на почве из пестрого песчаника | Сосновый лес на супесчаной почве | |
| 1 048 660 | 860 000 | 11 750 | |
| 149 800 | 168 666 | 11 750 | |
| 29 760 | 47 225 | 10 770 | |
| 11 980 | 14 708 | 3 525 | |
| 4 460 | 8 580 | 1 566 | |
| 2 630 | 4 272 | ||
| 1 488 | 2 471 | ||
| 1 018 | 1 735 | ||
| 1 398 | |||
| 1 057 | |||
| – | |||
| – | |||
| 145-150 | – |
|
|
|
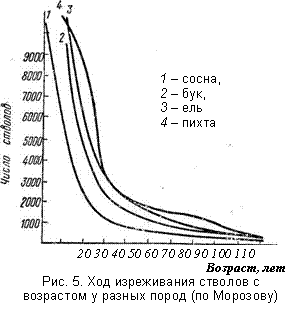
| Количество отмерших деревьев бука за 100 лет (от 10 до 110 лет) составило более 1 млн. на богатых почвах и более 850 000 на бедных, а для сосны – более 11000, что связано с малым числом стволов этой породы уже в десятилетнем возрасте. Сосна весьма свето-любива, поэтому уже к 10 годам у нее произошел значительный отпад. В результате за сто лет сохраняется одно дерево бука из 1800 на богатых почвах и из 950 – на более бедных, и одно дерево сосны из 28. На рис. 5 тоже показано, что гибель более светолюбивой породы (сосна) происходит быстрее, чем теневыносливых (бук, ель, пихта). Таким образом, различия в темпах изреживания у древостоя объясняются: 1) различным светолюбием (теневыносливостью); 2) увеличением темпа роста в хороших условиях и, как следствие, быстрым возрастанием его потребностей в экологических ресурсах, отчего конкуренция между видами становится все более напряженной. |

| Конкуренция внутри вида, значительно более интенсивна, чем между особями разных видов, но, в этом случае происходит дифференциация особей по высоте. В лесу деревья одного вида можно распределить по классам Крафта (рис. 6). Первый класс объединяет деревья хорошо развитые, поднимающиеся выше других – исключительно гос-подствующие, второй класс – господствующие, третий – согосподствующие, с развитыми, несколько сдавленными с боков, четвертый – заглушённые деревья, пятый – деревья угнетенные, отмирающие или отмершие. Сходная картина уменьшения количества экземпляров растений (на этот раз в течение одного сезона) и дифференциации по высоте наблюдается и в фитоценозах, образованных однолетними растениями, например солеросом травянистым (Salicornia herbacea). |
б) Экологический и фитоценотический оптимумы
|
|
|
Каждый вид имеет свою оптимальную плотность. Оптимальной плотностью называют те пределы плотности, которые обеспечивают наилучшее воспроизведение вида и его наибольшую устойчивость.
ПРИМЕРЫ. Для деревьев в рединах оптимальная плотность весьма невелика, они растут поодиночке на значительном расстоянии друг от друга, зато для лесообразующих пород она гораздо выше, а для болотных сфагновых мхов (Sphagnum) чрезвычайно высока.
Величина оптимальной площади и реакция на загущение зависит от тех условий, в которых проходила эволюция вида: одни виды развивались в условиях высокой плотности популяции, другие – в условиях низкой плотности; в одних случаях плотность была постоянной, в других – непрерывно меняющейся. Виды, эволюционировавшие в условиях постоянной плотности, резко реагируют на повышение плотности за пределы оптимальной замедлением роста; виды, развившиеся в условиях непрерывно менявшейся плотности, реагируют на изменение плотности за пределы оптимума слабо.
Каждый вид имеет два оптимума развития: экологический, сказывающийся на размерах особей вида, и фитоценотический, характеризующийся наиболее высокой ролью данного вида в фитоценозе, выражающейся в его обилии и степени проективного покрытия. Эти оптимумы и ареалы могут не совпадать. В природе чаще встречается фитоценотический оптимум, а экологический можно выявить, искусственно создавая растениям разные условия.
ПРИМЕРЫ. Многие галофиты лучше развиваются не на засоленных почвах, где они образуют сообщества, а на влажных почвах с незначительным содержанием солей. Многие растения скал с ксероморфным обликом свой экологический оптимум имеют на лугах.
Несовпадение экологического и фитоценотического оптимумов – результат борьбы за существование между растениями. В ряде случаев в процессе борьбы за существование растения оттесняются в экстремальные условия из более благоприятных для них фитоценозов.
ПРИМЕРЫ. Пихта белокорая и ель аянская не потому растут в более высоких горных поясах, что там лучше условия, а потому, что туда их вытесняют ель корейская, кедр и пихта цельнолиствная. Так же и светлолюбивые осина и береза уступают свои более благоприятные экотопы темнохвойным породам. Точно также травами из пойменных местообитаний вытесняются мхи и кустарнички.
|
|
|
в) Перенаселение вида
Для характеристики плотности вида существует такое понятие, как перенаселение. Рассматривают несколько видов перенаселения: абсолютное, относительное, возрастное, условное и локальное.
Под абсолютным перенаселением понимают такие условия загущения, при которых неизбежно наступает массовая гибель, носящая общий характер. (сверх густой посев – семена заделываются сплошным слоем или в два-три слоя), при котором, при условии очень дружных одновременных всходов на больших делянках, погибают все растения, кроме крайних).
Под относительным перенаселением понимают такие условия загущения, при которых гибель растений более или менее повышена, чем при густоте оптимальной для вида. При этом гибель растений избирательна, Действие отбора мягче, чем в случае абсолютного перенаселения.
Под возрастным перенаселением понимают перенаселение, возникающее с возрастом в результате неравномерного роста корневых систем (например, у корнеплодов) или надземных частей растений (у деревьев).
Условно перенаселенными называют сильно загущенные фитоценозы, в которых острота взаимоотношений между растениями снижена временной задержкой их роста до такой степени, что изреживание иногда полностью прекращается. Так, многие растения очень долго остаются в ювенильном (юношеском) состоянии, сохраняя весьма высокую выживаемость. Стоит вынудить растения к активному росту, как наступает настоящее перенаселение. Например, сильно угнетенные особи древесных пород под пологом густого леса имеет вид подроста.
Локальным перенаселением называют случаи перенаселения в гнездовых насаждениях очень высокой плотности и малой площади, в которых благодаря малой площади гнезда выживание каждой отдельной особи определяется не положением этой особи в гнезде, а ее особенностями, иными словами, гибель здесь носит избирательный характер.
Каково же значение явлений перенаселения для борьбы за существование и, следовательно, для процесса эволюции?
|
|
|
Перенаселение может иметь место в одних случаях и в одни периоды жизни растений и отсутствует в других случаях и в другие периоды жизни растений. В зависимости от степени перенаселения и от особенностей организмов оно способно как ускорять, так и замедлять процесс эволюции. При небольших степенях перенаселения оно вызывает дифференциацию особей и тем ускоряет процесс эволюции; при значительных степенях оно может вызвать обеднение популяции, уменьшение плодовитости и в результате – замедление эволюционного процесса. Перенаселение замедляет и ускоряет процесс естественного отбора, но не служит ему препятствием и не является непременным условием отбора, так как отбор может идти и без перенаселения.
Мы знаем, что для двух наиболее крупных групп органического мира – животных и растений – значение перенаселения неодинаково: оно играет гораздо большую роль в растительном мире, так как подвижность животных позволяет им в ряде случаев уйти от перенаселения.
Для разных систематических и экологических групп растений перенаселение играет не одну и ту же роль. Развитие большего, чем может впоследствии выжить, количества проростков и молодых растений обеспечивает виду господство в фитоценозе. Если бы проростки вида, преобладающего в фитоценозе, были единичными, то в массах развились бы проростки другого вида, и этот другой вид мог бы стать доминирующим в фитоценозе. Господствующий вид обычно дает большое количество проростков, но совершенно естественно, что лишь небольшая часть достигает зрелости. Значит, гибель большого количества молодых растений в данном случае неизбежна, именно она обеспечивает процветание вида и сохранение его положения в фитоценозе. Кроме молодых растеньиц погибает большое количество диаспор – зачатков растений (семена, плоды, споры) – еще до начала их развития (поедаются животными, погибают в. неблагоприятных условиях и т. д.). Таким образом, огромное количество диаспор, образуемых растениями, обеспечивает не только господство, но зачастую и само существование вида.
Внутривидовая конкуренция всегда более ожесточенна, чем межвидовая, поскольку особи одного вида более сходны друг с другом и предъявляют более сходные требования к среде, чем особи разных видов. Однако в природе, по-видимому, все сложнее. Так, при воспитании двух видов в чистых по севах и в смешанных (причем суммарное число особей на единицу площади в смешанном посеве равно числу особей на единицу площади в чистых посевах обоих видов) наблюдаются три типа взаимоотношений (Сукачев, 1953).
1. При совместном посеве оба вида развиваются лучше, чем каждый из них в одновидовом посеве. В этом случае межвидовая борьба оказывается слабее внутривидовой, что соответствует точке зрения Ч. Дарвина.
2. Из двух видов один лучше чувствует себя в смеси, чем в чистом посеве, а второй – хуже в смеси и лучше в чистом посеве. В этом случае для одного из видов межвидовая борьба оказывается более суровой, чем внутривидовая, а для другого – наоборот. Причины этого различны: выделение одним из видов колинов, вредных для особей другого вида, различие экологических особенностей видов, влияние продуктов разложения отмерших остатков одного вида на другой, различия в строении корневой системы и других особенностей.
3. Оба вида чувствуют себя в смеси хуже, чем в одновидовых посевах. В этом случае для обоих видов внутривидовая борьба менее сурова, чем межвидовая. Этот случай очень редок.
Следует иметь в виду, что отношения между парой каких-либо видов зависят от условий опыта: состава питательной среды, исходной численности растений, условий освещения, температурного режима и других причин.
|
|
|
|
Дата добавления: 2014-01-03; Просмотров: 833; Нарушение авторских прав?; Мы поможем в написании вашей работы!