КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Регистрация импульсной активности нервных клеток
|
|
|
|
Пейсмекер
Один из удивительных видов электрической активности нейронов, регистрируемой внутриклеточным микроэлектродом, – это пейсмекерные потенциалы. А. Арванитаки и Н. Халазонитис [Arvanitaki, Chalazonitis, 1955] впервые описали осциллирующие потенциалы нервной клетки, не связанные с поступлением к ней синаптических влияний. Эти колебания в ряде случаев могут приобретать такой размах, что превышают критический уровень потенциала, необходимый для активации механизма электрической возбудимости. Наличие в соме клетки таких волн мембранного потенциала было обнаружено на нейронах моллюсков. Они были расценены как проявление спонтанной, или ауторитмической активности, имеющей эндогенное происхождение [Alving, 1968].
Аналогичные ритмические колебания были затем описаны и во многих других типах нейронов. Способность к длительной ритмической активности сохраняется у некоторых клеток в течение длительного времени после полного их выделения [Chen et al., 1971; Греченко, Соколов, 1986]. Следовательно, в ее основе действительно лежат эндогенные процессы, приводящие к периодическому изменению ионной проницаемости поверхностной мембраны. Важную роль играют изменения ионной проницаемости мембраны под действием некоторых цитоплазматических факторов, например системы обмена циклических нуклеотидов. Изменения активности этой системы при действии на соматическую мембрану некоторых гормонов или других внесинаптических химических влияний могут модулировать ритмическую активность клетки (эндогенная модуляция).
Запускать генерацию колебаний мембранного потенциала могут синаптические и внесинаптические влияния. Л. Тауц и Г.М. Гершенфельд [Tauc, Gerschenfeld, 1960] обнаружили, что соматическая мембрана нейронов моллюсков, не имеющая на своей поверхности синаптических окончаний, обладает высокой чувствительностью к медиаторным веществам и, следовательно, имеет молекулярные хемоуправляемые структуры, свойственные постсинаптической мембране. Наличие внесинаптической рецепции показывает возможность модуляции пейсмекерной активности диффузным действием выделяющихся медиаторных веществ.
|
|
|
Сложившаяся концепция о двух типах мембранных структур – электровозбудимой и электроневозбудимой, но химически возбудимой, заложила основу представлений о нейроне как пороговом устройстве, обладающем свойством суммации возбуждающих и тормозных синаптических потенциалов. Принципиально новое, что вносит эндогенный пейсмекерный потенциал в функционирование нейрона, заключается в следующем: пейсмекерный потенциал превращает нейрон из сумматора синаптических потенциалов в генератор. Представление о нейроне как управляемом генераторе заставляет по-новому взглянуть на организацию многих функций нейрона.
Пейсмекерными потенциалами в собственном смысле этого слова называют близкие к синусоидальным колебания частотой 0,1–10 Гц и амплитудой 5–10 мВ. Именно эта категория эндогенных потенциалов, связанных с активным транспортом ионов, образует механизм внутреннего генератора нейрона, обеспечивающего периодическое достижение порога генерации ПД в отсутствие внешнего источника возбуждения. В самом общем виде нейрон состоит из электровозбудимой мембраны, химически возбудимой мембраны и локуса генерации пейсмекерной активности. Именно пейсмекерный потенциал, взаимодействующий с хемовозбудимой и электровозбудимой мембраной, делает нейрон устройством со «встроенным» управляемым генератором [Bullock, 1984].
Если локальный потенциал является частным случаем механизма генерации ПД, то пейсмекерный потенциал принадлежит к особому классу потенциалов – электрогенному эффекту активного транспорта ионов. Особенности ионных механизмов электрической возбудимости соматической мембраны лежат в основе важных свойств нервной клетки, в первую очередь ее способности генерировать ритмические разряды нервных импульсов. Электрогенный эффект активного транспорта возникает в результате несбалансированного переноса ионов в разных направлениях. Широко известен гиперполяризационный постоянный потенциал как результат активного вывода ионов натрия, суммирующийся потенциалом Нернста [Ходоров, 1974]. Дополнительное включение активного насоса ионов натрия создает фазичекие медленные волны гиперполяризации (негативные отклонения от уровня мембранного потенциала покоя), обычно возникающие вслед за высокочастотной группой ПД, которая приводит к избыточному накоплению натрия в нейроне.
|
|
|
Несомненно, что некоторые из компонентов механизма электрической возбудимости соматической мембраны, а именно электроуправляемые кальциевые каналы, вместе с тем являются фактором, сопрягающим мембранную активность с цитоплазматическими процессами, в частности с процессами протоплазматического транспорта и нервной трофики. Детальное выяснение этого важного вопроса требует дальнейшего экспериментального изучения.
Пейсмекерный механизм, являясь эндогенным по происхождению, может активироваться и инактивироваться на длительное время в результате афферентных воздействий на нейрон. Пластические реакции нейрона могут обеспечиваться изменениями эффективности синаптической передачи и возбудимости пейсмекерного механизма (Соколов, Тавкхелидзе, 1975).
Пейсмекерный потенциал является компактным способом передачи внутринейронной генетической информации. Приводя к генерации ПД, он обеспечивает возможность выхода эндогенных сигналов на другие нейроны, в том числе и эффекторные, обеспечивающие реакцию [Bullock, 1984]. Тот факт, что генетическая программа включает звено управления пейсмекерным потенциалом, позволяет нейрону реализовывать последовательность своих генетических программ. Наконец пейсмекерный потенциал в той или иной степени может подвергаться синаптическим влияниям. Этот путь позволяет интегрировать генетические программы с текущей активностью, обеспечивая гибкое управление последовательными программами. Пластические изменения пейсмекерного потенциала еще больше расширяют возможность приспособления наследственно фиксированных форм к потребностям организма. Пластические изменения развиваются в этом случае не в геноме, а на пути выхода наследственной программы на реализацию (на уровне генерации ПД).
|
|
|
Глава 2 МЕТОДЫ ПСИХОФИЗИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ
Физиологические процессы, как правило, скрыты от внешнего наблюдения, поэтому они длительное время оставались вне области интересов психологов, занимавшихся в основном исследованием доступных для прямого наблюдения проявлений поведения человека. Однако многие модели психической деятельности человека носили бы чисто умозрительный характер, а психология оставалась бы «безмозглой», если бы психологи не заинтересовались нейрофизиологическими процессами, лежащими в основе исследуемой ими реальности [Швырков, 1995].
С другой стороны, в нейрофизиологии постоянно возникала потребность описать организацию физиологических процессов в терминах, определяемых в психологических концепциях и теориях. Происходило и происходит взаимное обогащение двух наук о человеке как теоретическими разработками, так и экспериментальными методами (см. Предисловие). Что же дает изучение физиологических показателей работы нервной системы? Во-первых, в силу своей объективности физиологические показатели становятся надежными элементами, используемыми при описании изучаемого поведения. Во-вторых, они позволяют экспериментаторам включить в сферу своих исследований скрытые для прямого наблюдения проявления активности организма, лежащие в основе поведения. И, как оптимистично заявил Ж. Пайяр: «Помимо более полного объективного описания явлений, в основе обращения к физиологическим показателям лежит смелое устремление, которое продолжает направлять усилия современных психологов объяснить психологические явления на органической основе» [1970, с.10].
|
|
|
В психофизиологии основными методами регистрации физиологических процессов являются электрофизиологические методы. В физиологической активности клеток, тканей и органов особое место занимает электрическая составляющая. Электрические потенциалы отражают физико-химические следствия обмена веществ, сопровождающие все основные жизненные процессы, и поэтому являются исключительно надежными, универсальными и точными показателями течения любых физиологических процессов [Коган, 1969]. Надежность электрических показателей по сравнению с другими, по мнению А.Б. Когана, особенно демонстративна, «когда они оказываются единственным средством обнаружения деятельности» [там же, с.13]. Единообразие потенциалов действия в нервной клетке, нервном волокне, мышечной клетке как у человека, так и у животных говорит об универсальности этих показателей. Точность электрических показателей, т.е. их временное и динамическое соответствие физиологическим процессам, основана на быстрых физико-химических механизмах генерации потенциалов, являющихся неотъемлемым компонентом физиологических процессов в нервной или мышечной структуре.
К перечисленным преимуществам электрических показателей физиологической активности следует добавить и неоспоримые технические удобства их регистрации: помимо специальных электродов, для этого достаточно универсального усилителя биопотенциалов, который скоммутирован с компьютером, имеющим соответствующее программное обеспечение. И, что важно для психофизиологии, большую часть этих показателей можно регистрировать, никак не вмешиваясь в изучаемые процессы и не травмируя объект исследования. К наиболее широко используемым методам относятся регистрация импульсной активности нервных клеток, регистрация электрической активности кожи, электроэнцефалография, электроокулография, электромиография и электрокардиография. В последнее время в психофизиологию внедряется новый метод регистрации электрической активности мозга – магнитоэнцефалография и изотопный метод (позитронноэмиссионная номография).
Изучение активности нервных клеток, или нейронов, как целостных морфологических и функциональных единиц нервной системы, безусловно, остается базовым направлением в психофизиологии. Одним из показателей активности нейронов являются потенциалы действия – электрические импульсы длительностью несколько мс и амплитудой до нескольких мВ. Современные технические возможности позволяют регистрировать импульсную активность нейронов у животных в свободном поведении и, таким образом, сопоставлять эту активность с различными поведенческими показателями. В редких случаях в условиях нейрохирургических операций исследователям удается зарегистрировать импульсную активность нейронов у человека.
Поскольку нейроны имеют небольшие размеры (несколько десятков микрон), то и регистрация их активности осуществляется с помощью подводимых вплотную к ним специальных отводящих микроэлектродов. Свое название они получили потому, что диаметр их регистрирующей поверхности составляет около одного микрона. Микроэлектроды бывают металлическими и стеклянными. Металлический микроэлектрод представляет собой стержень из специальной высокоомной изолированной проволоки со специальным способом заточенным регистрирующим кончиком. Стеклянный микроэлектрод – пирексовая тонкая трубочка (диаметр около 1 мм) с тонким незапаянным кончиком, заполненная раствором электролита. Электрод фиксируется в специальном микроманипуляторе, укрепленном на черепе животного, и коммутируется с усилителем. С помощью микроманипулятора электрод через отверствие в черепе пошагово вводят в мозг. Длина шага составляет несколько микрон, что позволяет подвести регистрирующий кончик электрода очень близко к нейрону, не повреждая его (рис. 2.1 А). Подведение электрода к нейрону осуществляется либо вручную, и в этом случае животное должно находиться в состоянии покоя, либо автоматически на любом этапе поведения животного. Усиленный сигнал поступает на монитор и записывается на магнитную ленту или в память ЭВМ. При «подходе» кончика электрода к активному нейрону экспериментатор видит на мониторе появление импульсов, амплитуда которых при дальнейшем осторожном продвижении электрода постепенно увеличивается. Когда амплитуда импульсов начинает значительно превосходить фоновую активность мозга, электрод больше не подводят, чтобы исключить возможность повреждения мембраны нейрона. Пример импульсной активности нейрона, зарегистрированной у кролика, находящегося в условиях свободного поведения, представлен на рис. 2.1 В.
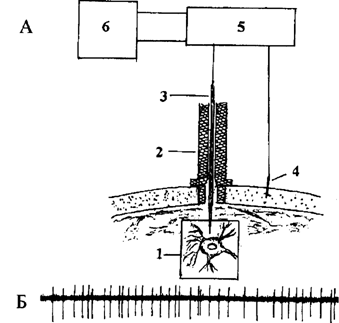
Рис. 2.1. А – принципиальная схема регистрации импульсной активности нейрона:
1 - нейрон (увеличен) и кончик отводящего электрода;
2 – микроманипулятор (в разрезе);
3 – микроэлектрод с отводящим проводом;
4 – индифферентный электрод;
5 – усилитель;
6 – монитор и записывающее устройство
Б – пример записи импульсной активности нейрона (нейронограмма)
|
|
|
|
|
Дата добавления: 2017-01-14; Просмотров: 887; Нарушение авторских прав?; Мы поможем в написании вашей работы!