КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Биотический потенциал
|
|
|
|
Динамика популяций
Вопрос № 24
Динамика популяции – это процессы изменения ее основных биологических характеристик во времени. Можно выделить два основных типа кривых роста численности популяции.
Любая популяция теоретически способна к неограниченному росту численности, если ее не лимитируют факторы внешней среды. В таком гипотетическом случае скорость роста популяции будет зависеть только от величины биотического потенциала, свойственного виду. Понятие биотического потенциала введено в экологию в 1928 г. Р.Чепменом. Этот показатель отражает теоретический максимум потомков от одной пары (или одной особи) за единицу времени, например за год или за весь жизненный цикл.При расчетах его чаще всего выражают коэффициентом г и вычисляют как максимально возможный прирост популяции?N за отрезок времени? t, отнесенный к одной особи, при начальной численности популяции N0:
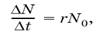 откуда
откуда 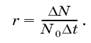
 1. Экспоненциальный рост численности (J -образная кривая) (рис. 9):
1. Экспоненциальный рост численности (J -образная кривая) (рис. 9):
N t = N 0 e rt, где N t – численность популяции в момент времени t, N 0 – численность популяции в начальный момент времени t 0, е – основание натурального логарифма, r – показатель, характеризующий темп размножения особей в данной популяции (удельная скорость изменения численности). Если r > 0, то численность популяции растет, при r < 0 – сокращается. Экспоненциальный рост возможен лишь при отсутствии лимитирующих факторов. Такой рост в природе не происходит, либо происходит в течение очень непродолжительного времени (например, популяции одноклеточных организмов, водорослей, мелких ракообразных при благоприятных условиях размножаются по экспоненциальному закону). Это рост численности особей в неизменяющихся условиях.
|
|
|
Рис. 122. Реальная (1) и теоретическая (2) кривая роста популяции парамеций
Воздействие экологических факторов на скорость роста популяции может довести численность популяции до стабильной (r = 0) или уменьшить ее, т.е. экспоненциальный рост замедляется или останавливается. J -образная кривая превращается в S -образную.
2. Логистическая кривая роста (S -образная кривая) (рис. 10): скорость роста популяции линейно снижается по мере роста численности до 0 при некоторой предельной численности К (К – максимальное число особей, способных жить в рассматриваемой среде). При N 0 r = max, а при N = K r = 0. В дифференциальной форме логистическое уравнение выглядит следующим образом: d N /d t = r max N (K – N)/ K
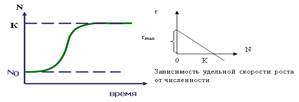
В природе биотический потенциал популяции никогда не реализуется полностью. Его величина обычно складывается как разность между рождаемостью и смертностью в популяциях: r = b – d, где b – число родившихся, а d – число погибших особей в популяции за один и тот же период времени.
Рождаемость – это число новых особей, появляющихся в популяции за единицу времени в расчете на определенное число ее членов.
абсолютная-(B) характеризуется общим числом особей, появляющихся в единицу времени в данной популяции, и определяется по формуле: B = A Nn/ A t, где A Nn - количество особей, появившихся в популяции за время A t, A t, - анализируемый промежуток времени. общее число родившихся особей(на 16тыс голов2 тыс оленят)
удельная-(b) характеризу-ется средним числом особей, появившихся в единицу времени в расчете на одну особь популяции, и определяется по формуле: b = ANn/(At x No) = B/No, где B - абсолютная рождаемость, No - первоначальная численность популяции.среднее значение 0,125 т.е один новорожденный на 8 членов популяции за год
Величина рождаемости зависит от многих причин. Большое значение имеет доля особей, способных в данный период к размножению, что определяется соотношением полов и возрастных групп. Важна также частота последовательности генераций.
|
|
|
1. Моновольтинные - одна генерация в год.
2. Поливольтинные -несколько генерации в год(тля)
По числу периодов размножения в течение жизни различают
1. Моноцикличность, или однократное размножение, свойственна обычно видам с короткой продолжительностью жизни в половозрелом состоянии (лососевые рыбы, поденки, майские жуки и многие другие насекомые).
2 Полицикличность характеризуется повторным размножением особей и присуща большинству позвоночных животных и ряду беспозвоночных, например ракообразным.
У растений выделяют монокарпические и поликарпические виды, т. е. с однократным и многократным размножением в течение жизни.
Смертность – величина обратная рождаемости, число особей, погибших в популяции за единицу времени от всех причин. Различают минимальную (минимально возможную) и фактическую смертность. Также различают абсолютную и удельную смертность, которые вычисляются так же, как и рождаемость.
Различия в биотическом потенциале видов зависят от их размеров, систематической принадлежности и других причин, но при прочих равных условиях связаны зависимостью со смертностью в популяциях. Эта закономерность, подмеченная еще Ч. Дарвином, была обоснована в трудах академика И. И. Шмальгаузена в 40‑е годы прошлого столетия. Если вид подвергается в природе массовой неизбирательной элиминации, т. е. гибели от многочисленных врагов, избежать которых он бессилен, или подавляется другими экстремальными обстоятельствами, то единственным направлением отбора становится повышение размножаемости. В этом случае увеличивается вероятность случайного сохранения потомства и вид избегает вымирания. При неизбирательной элиминации различия между особями не имеют значения для их выживания, поскольку мощность воздействия губительных факторов слишком высока. При избирательной элиминации, когда смертность во многом определяется различиями между особями, отбор совершенствует разные формы морфофизиологических адаптаций, повышающих сопротивляемость вида влиянию неблагоприятных условий. Таким образом, высокий биотический потенциал – эволюционный ответ вида на пресс неблагоприятных для него воздействий среды, вызывающих высокую смертность.
|
|
|
В конце 60‑х годов эта идея возродилась в концепции К– и г ‑отбора, выдвинутой американскими экологами Р. Макартуром и Э. Уилсоном. Они предложили различать две основные стратегии размножения организмов, обеспечивающие выживание в разных условиях, обозначив их через коэффициенты, входящие в уравнение роста популяций. При r‑стратегии отбор идет на высокую плодовитость, оборачиваемость поколений, способность к быстрому расселению, что позволяет видам быстро восстанавливать численность после резкого ее снижения. При К‑стратегии отбор совершенствует разные формы заботы о потомстве, что позволяет снизить плодовитость. Одновременно увеличивается продолжительность жизненных циклов и совершенствуются механизмы устойчивого поддержания численности в биоценозах. Естественно, что между крайними формами имеются все промежуточные варианты. Элементы К– и г ‑стратегий выживания прослеживаются во всех систематических группах организмов. Даже в пределах вида в популяциях, обитающих в разных условиях, усиливаются те или иные направления отбора.
На практике, когда необходимо проанализировать ход смертности в популяциях, составляют таблицы выживания (см. далее), где для каждой возрастной группы указываются эмпирически полученные данные, характеризующие гибель особей данного возраста.
На основе таких таблиц составляют кривые выживания, позволяющие прогнозировать в сходных условиях состояние очередных генераций.
Зависимость числа выживших особей от времени (или возраста) называют кривыми выживания. Кривые выживания подразделяют на три основных типа (рис. 8):

1. Кривая I типа – сильно выпуклая кривая, характерна для видов, у которых на протяжении всей жизни смертность мала и резко возрастает в конце жизни (дрозофила, человек).
2. Кривая III типа – сильно вогнутая кривая, характерна для видов с высокой смертностью в начальный период жизни (устрица).
3. Кривая II типа (диагональная) характерна для видов, у которых смертность остается примерно постоянной в течение всей жизни (гидра, рыбы, птицы).
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 2679; Нарушение авторских прав?; Мы поможем в написании вашей работы!