КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Белки-репрессоры бактерий связываются вблизи промоторов и подавляют транскрипцию определенных
|
|
|
|
10-8
10-5
10.1.8. Один-единственный главный белок-регулятор может превратить фибробласт в миобласт [7]
Клетка скелетной мышцы млекопитающих обычно очень велика и содержит много ядер. Она образуется в результате слияния многих клеток-предшественников, называемых миобластами (см. разд. 17.6.1). Зрелая мышечная клетка отличается от других клеток большим числом присущих только ей белков, включая специфические типы актина, миозина, тропомиозина и тропонина (входящих в состав сократительного аппарата), креатинфосфокиназу (обеспечивающую специализированный метаболизм мышечной клетки) и ацетилхолиновые рецепторы (необходимые для того, чтобы мембрана была чувствительна к нейростимуляции). В пролиферирующих миобластах такие специфичные для мышц белки и соответствующие им мРНК отсутствуют либо обнаруживаются в очень небольших количествах. По мере слияния миобластов в образующихся клетках одновременно увеличивается концентрация мышечно-специфичных белков и их мРНК. Следовательно, контроль за экспрессией соответствующих генов осуществляется на уровне транскрипции. Уровень синтеза многих специфичных для мышцы белков возрастает по меньшей мере в 500 раз. С помощью двумерного электрофореза в полиакриламидном геле показано, что одновременно меняется концентрация многих других белков: некоторые из них перестают синтезироваться, продукция других достигает максимума и затем падает, у части белков происходит сдвиг с одного уровня синтеза на другой и так далее.
Если в культивируемые фибробласты кожи (клетки, которые в норме никогда не экспрессируют мышечно-специфичные гены) ввести ген регуляторного белка из миобласта (myo D1), такие клетки приобретают способность к слиянию и другие свойства мышечных клеток (рис. 10-10).
|
|
|
Очевидно myo D1 является главным регуляторным белком, который в норме и определяет «миобласт».
Белок myo D1 сосредоточен в клеточном ядре. Исходя из нуклеотидной последовательности ДНК, была определена последовательность аминокислот в его молекуле. Изучение белка myo D1 показало, что он

Рис. 10-10. Иммунофлуоресцентная микрофотография фибробластов кожи куриного эмбриона, превратившихся в клетки мышц при экспрессии в них гена myoD1, которая была вызвана искусственным путем. Эти фибробласты выращивали в культуре и за три дня до съемки трансформировали рекомбинантной плазмидной ДНК. В состав этой плазмиды входили последовательности, кодирующие белок myoD, которые соединены с промотором/энхансером вирусного происхождения, активным в куриных эмбрионах. Несколько процентов фибробластов включало ДНК и продуцировало белок myoD. Эти клетки слились с образованием удлиненных миофибрилл, которые на этой фотографии помечены антителами, выявляющими белок, специфичный для мышц. В контрольной культуре, трансфицированной другой плазмидой, мышечные клетки отсутствуют. (С любезного разрешения Stephen Tapscott, Andrew Lassar, Robert Davis и Harold Weintraub.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
связывается с регуляторними участками генов, специфичных для мышц. Свойства этого замечательного белка подтверждает предположение о том, что главные регуляторные белки способны детерминировать превращение клеток.
Заключение
У многоклеточных организмов дифференцировка клеток происходит в результате экспрессии разных генов одного и того же генома, хотя типы клеток на удивление мало отличаются друг от друга по содержанию белков. Экспрессия большинства генов контролируется на уровне транскрипции, что не исключает существенной роли посттранскрипционного контроля. Контроль на уровне транскрипции зависит от регуляторных белков, связывающихся с определенными последовательностями ДНК. В результате присоединения таких белков соответствующие гены либо включаются (позитивный контроль) либо выключаются (негативный контроль). Гены высших эукариот обычно регулируются путем комбинационного воздействия нескольких белков-регуляторов, осуществляющих позитивный и негативный контроль. Главные регуляторные белки играют в системе регуляции активности генов особую роль благодаря тому, что они влияют на активность сразу многих генов: например, экспрессия гена mуо D1 может превратить фибробласт в миобласт.
|
|
|
10.2. Контроль инициации транскрипции [8]
Всего лишь 40 лет назад мысль о том, что ген можно включать или выключать, казалась абсурдной. Гипотеза, сыгравшая такую важную роль в понимании работы клеток, была выдвинута на основании изучения Е. coli, растущей на смеси глюкозы и лактозы (дисахарид). Если бактерии предоставляли выбор источника углерода, она сначала использовала всю глюкозу и лишь затем начинала метаболизировать лактозу. Переключение на лактозу сопровождалось остановкой роста, в течение которой синтезировался фермент β-галактозидаза, гидролизирующий лактозу до глюкозы и галактозы. Выделение и характеристика мутантных бактерий, обладающих определенными дефектами в регуляции такого переключения, дало толчок биохимическим исследованиям, которые в 1966 г. привели к идентификации и выделению белка-penpeccopa лактозного оперона.
В результате биохимического и генетического изучения белка-репрессора лактозного оперона, репрессора бактериофага лямбда, а также других регуляторных белков у бактерий была сформулирована общая модель регуляции транскрипции у прокариот. Предполагалось, что сайт-
специфические белки либо ингибируют, либо стимулируют транскрипцию какого-либо гена, присоединяясь к ДНК рядом с промотором-участком, с которого РНК-полимераза начинает синтез РНК. Считали, что изменение в положении таких регуляторных белков по отношению к ДНК (связывание или отсоединение) включает и выключает гены.
|
|
|
В течение многих лет оставалось неизвестным, можно ли применять эту модель контроля генетической активности к клеткам эукариот.
Известно, что ДНК эукариотической клетки упакована с помощью гистоновых белков в нуклеосомы, масса которых равна массе ДНК. Присутствие нуклеосом предполагает существование каких-то иных путей регуляции генов. Об этом же свидетельствует и тот факт, что у эукариот регуляторные белки часто связываются в участках, удаленных на тысячи нуклеотидных пар от промотора, на который они
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
действуют. Возможности изучения механизма регуляции генов эукариот были ограничены главным образом из-за того, что большинство белков-
регуляторов присутствует в клетке в очень малых количествах (примерно один из 50000 молекул белка). Только в 1983 г. удалось охарактеризовать два необычно многочисленных (и, возможно, нетипичных) белка -регулятора - большой Т-антиген вируса SV40 и фактор транскрипции TF1IIA гена 5S-pPHK.
В последнее время ситуация в корне изменилась. Благодаря новым методам генной инженерии для биохимического и генетического анализа стало доступным большое число белков-регуляторов эукариот. Кроме того, у бактерий были выявлены белки-регуляторы, осуществляющие свое действие на расстоянии. Данный раздел посвящен тому общему, что объединяет механизмы регуляции транскрипции генов у прокариот и у эукариот. Процессы, которые могут оказаться характерными только для эукариот, мы обсудим позднее в связи с проблемой дифференцировки клеток (см. разд. 10.3.8).
генов [9]
Хромосома Е, coli состоит из единственной кольцевой молекулы ДНК, насчитывающей ~ 4,7 х 10б нуклеотидных пар. В принципе такого количества достаточно для кодирования примерно 4000 различных белков, однако в конкретный момент времени в клетке синтезируется лишь определенная часть этих белков. Экспрессия многих генов E.coli зависит от внутриклеточного уровня специфических метаболитов.
|
|
|
Изучение генетического контроля утилизации лактозы в клетках Е. coli привело исследователей к выводу о существовании белка-
репрессора, который в отсутствие лактозы в среде выключает синтез β-галактозидазы. Выделение, очистка этого белка и использование его в опытах in vitro позволили расшифровать механизм такой регуляции. Оказалось, что репрессор ингибирует транскрипцию lac-оперона, связываясь с так называемым оператором - последовательностью ДНК длиной 21 нуклеотид, которая перекрывается с расположенным по соседству участком связывания РНК-полимеразы (промотором). До тех пор, пока репрессор
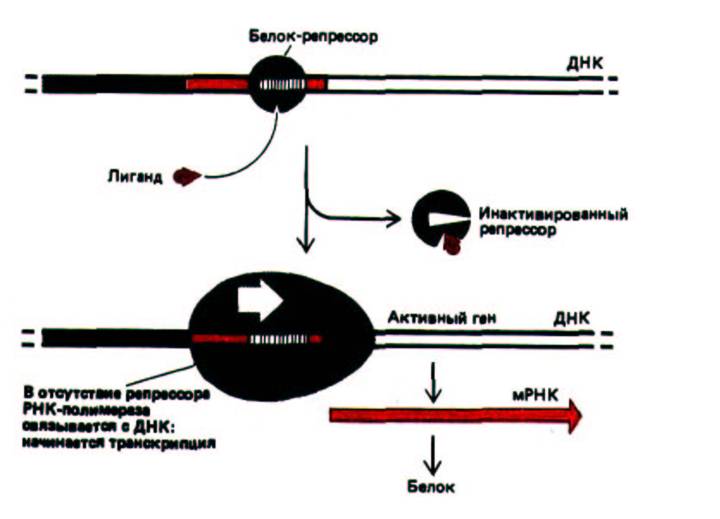
Рис. 10-11. Схема дерепрессии у бактерий. Небольшая специфическая молекула связывается с белком-репрессором, в результате чего происходит изменение его конформации и отделение от ДНК, при этом расположенные рядом гены могут начать транскрибироваться. В случае примера, описанного в тексте, аллолактоза связывается с белком-репрессором лактозного оперона, в результате чего синтезируется единственный транскрипт РНК, кодирующий три белка, которые участвуют в метаболизме лактозы (β-галактозидаза, галакто-зидпермеаза и галактозидацетилаза).
Поскольку три гена, кодирующие эти белки, расположены рядом друг с другом и их регуляция осуществляется строго координирование, кластер этих генов называют лактозным опероном.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
остается связанным с оператором, доступ РНК-полимеразы к ее участку связывания закрыт и транскрипция близлежащих областей ДНК не происходит (см. рис. 10-14).
Белок-реперессор лактозного оперона отвечает за то, чтобы уровень синтеза β-галактозидазы - фермента, необходимого для расщепления лактозы, соответствовал потребностям клетки. Лактозный репрессор в свою очередь находится под контролем небольшой углеводной молекулы аллолактозы, которая образуется в клетке в присутствии лактозы. Когда внутриклеточное содержание аллолактозы достигает достаточно высокого уровня, она, выполняя функцию аллостерического регулятора, индуцирует конформационные изменения в молекуле репрессора. При этом его связь с ДНК ослабляется настолько, что он отделяется от нее, освобождая промотор и давая возможность РНК-полимеразе транскрибировать прилежащие области ДНК. В этом случае говорят о дерепрессии гена. Такая система регуляции позволяет E.coli производить фермент, необходимый для расщепления лактозы, лишь тогда, когда это необходимо (рис. 10-11).
В настоящее время известно много других примеров специфической репрессии бактериальных генов. В каждом случае связывание белка-
репрессора с определенной последовательностью ДНК приводит к выключению гена. Процесс связывания всегда регулируется определенными сигнальными молекулами, аналогичными аллолактозе. Иногда, как в случае репрессора лактозного оперона, присутствие сигнальных молекул в клетке включает ген или единицу транскрипции, уменьшая сродство белка-репрессора к определенной последовательности ДНК. Но сигнальная молекула может использоваться и для выключения гена с помощью белка-репрессора. Например, аллостерическое изменение, вызванное связыванием сигнальной молекулы с репрессором, может повысить, а не понизить способность репрессора связываться с определенной последовательностью ДНК. Такой механизм действует в случае контроля активности пяти расположенных рядом генов, которые кодируют ферменты, необходимые для синтеза триптофана в клетках E.coli (trp-onepoн). Синтез единственной длинной молекулы мРНК, кодирующей эти пять белков, контролируется белком-репрессором, который «садится» на ДНК лишь в том случае, если он связан с триптофаном (сигнальная молекула, включающая этот оперон) (рис. 10-12).
В случае laс-оперона повышение концентрации сигнальных молекул

Рис. 10-12. Соединение триптофана с белком-репрессором триптофанового оперона изменяет конформацию репрессора.
Конформационные изменения дают возможность этому регуляторному белку тесно связываться со специфической последовательностью ДНК, и, таким образом, блокировать транскрипцию генов, которые кодируют белки, участвующие в синтезе триптофана (trp-оперон). Трехмерная структура этого бактериального белка (спираль-виток-спираль) определена с помощью метода рассеивания рентгеновских лучей и показана как в случае связывания триптофана, так и без него. Связывание триптофана приводит к увеличению расстояния между двумя узнающими спиралями (цветные цилиндры) в димере, что способствует образованию симметрично расположенных водородных связей, изображенных на схеме в виде цветных лучей. (По R. Zhang et al., Nature, 327: 591-597, 1987.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
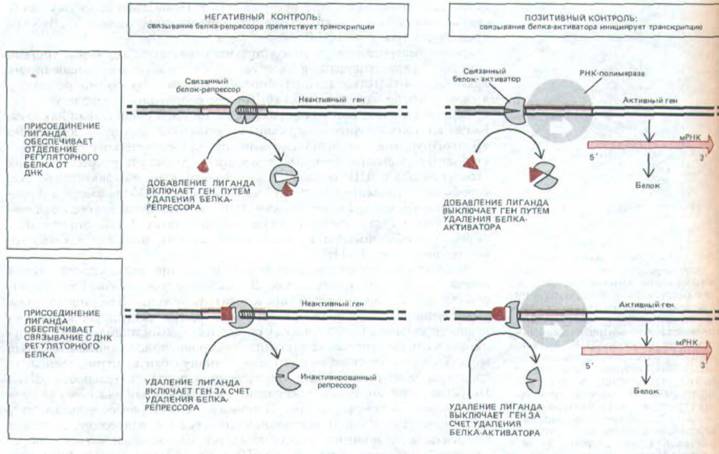
Рис. 10-13. Обобщенная схема различных механизмов, с помощью которых специфические регуляторные белки контролируют транскрипцию генов у прокариот. А. Негативный контроль; Б. Позитивный контроль. Обратите внимание, что лиганд-индуктор может включить ген, либо обеспечивая удаление белка-репрессора (слева вверху), либо способствуя присоединению к ДНК белка-активитора (справа внизу).
Аналогичным образом лиганд-ингибитор может выключить ген как путем удаления белка-активатора (справа вверху), так и путем связывания белка-репрессора с ДНК (слева внизу).
способствует освобождению ДНК от репрессора и активации транскрипции, а в случае trp-оперона вызывает связывание репрессора с ДНК и, таким образом, подавляет транскрипцию. Следует, однако, подчеркнуть, что оба описанных механизма в принципе одинаковы: и в том, и в другом случае транскрипция осуществляется в отсутствие регуляторного белка. Генетический контроль такого типа называется негативной регуляцией (рис. 10-
13, А).
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 1482; Нарушение авторских прав?; Мы поможем в написании вашей работы!