КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Продукты некоторых онкогенов - это аномальные каталитические рецепторы с нерегулируемой киназной
|
|
|
|
активностью [21]
Первая тирозиновая протеинкиназа была открыта в 1979 г. Это был не поверхностный клеточный рецептор, а внутриклеточный продукт вирусного онкогена - белок, названный рр60 v-src (разд. 13.4.2). Первым рецептором, у которого обнаружили тирозинкиназную активность (в 1982
г.), был рецептор для EGF. Несколькими годами позже выяснилось, что вирусный онкоген erbB кодирует урезанный вариант рецептора для EGF.
Этот урезанный белок потерял EGF-связывающий наружный домен, но сохранил внутриклеточный домен с тирозинкиназной активностью, и поэтому клетки с такими дефектными рецепторами ведут себя так, как будто на них постоянно действует сигнал к пролиферации. Позднее выяснилось, что онкоген пеи, активный в некоторых химически индуцированных опухолях нервной системы у крыс, кодирует аномальный рецептор, являющийся тирозиновой киназой, хотя природа лиганда (предположительно это ростовой фактор) для нормального рецептора не установлена. В этом случае аномальный и нормальный рецепторы различаются только по одному аминокислотному остатку в единственном трансмембранном сегменте белка. Такого изменения оказалось достаточно, чтобы сделать тирозиновую киназу постоянно активной. Эти исследования подчеркивают важную роль тирозиновых киназ в контроле клеточной пролиферации.
Между онкогенами и нормальными путями передачи сигнала к пролиферации было обнаружено немало и других связей. Онкоген sis, например, кодирует функционально активную субъединицу PDGF, а онкоген erbA - измененную форму рецептора тиреоидного гормона. Как мы будем более подробно обсуждать в гл. 13, исследование онкогенов открывает перспективный путь к выявлению и пониманию целого спектра механизмов, с помощью которых пролиферативные сигналы достигают своей цели.
|
|
|
Продукты примерно половины всех открытых до сих пор онкогенов - это протеинкиназы, фосфорилирующие белки-мишени по остаткам тирозина, серина или треонина. Это не удивительно, так как фосфорилирование играет важную роль в процессах передачи сигнала, запускаемых как каталитическими рецепторами, так и рецепторами, сопряженными с G-белками, и для его осуществления имеется весьма обширное семейство протеинкиназ. Уже известно более 70 протеинкиназ, и все они, видимо, происходят от общего предшественника, так как их каталитические домены гомологичны (рис. 12-25). Фактически сейчас уже возможно предсказать, будет ли белок киназой и если да, то какие остатки - серина, треонина или тирозина - он будет фосфорилировать, просто исходя из данных о его аминокислотной последовательности. В следующем разделе мы увидим, что два главных внутриклеточных посредника - сАМР и Са2+-реализуют многие свои эффекты, активируя протеинкиназы, специфичные в отношении серина и треонина.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
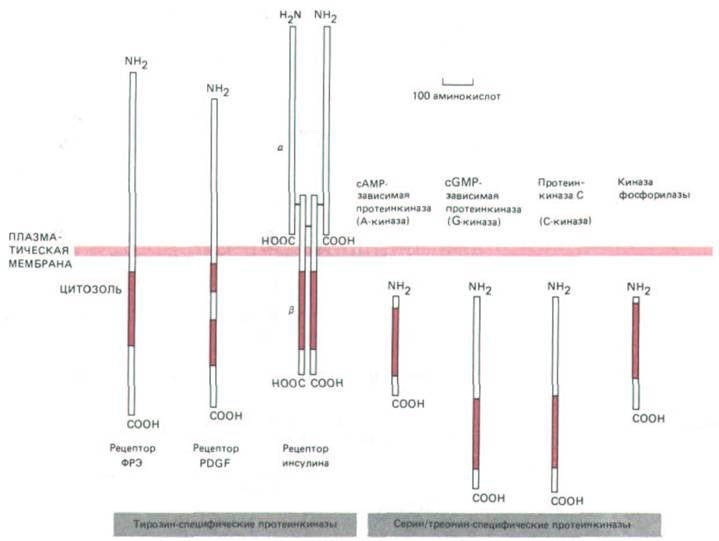
Рис. 12-25. Размеры и локализация каталитических доменов некоторых протеинкиназ, рассмотренных в этой главе. Во всех случаях каталитический домен (выделен цветом) состоит примерно из 250 аминокислотных остатков и имеет сходную аминокислотную последовательность; это позволяет предполагать происхождение их всех от общего предшественника. Три представленные здесь тирозин-
специфические киназы-трансмембранные белки-рецепторы, которые при связывании специфического внеклеточного лиганда активируются и фосфорилируют ряд белков внутри клетки (в том числе и самих себя) по остаткам тирозина. Обе цепи рецептора инсулина кодируются одним геном, продукт которого - белок-предшественник - расщепляется на две цепи, связанные дисульфидными мостиками. Внеклеточная часть рецептора PDGF, по-видимому, сложена в пять иммуноглобулиноподобных доменов - возможно, этот белок относится к суперсемейству иммуноглобулинов (разд. 18.6.20). Регуляторные субъединицы А-киназы (см. рис. 12-27) и киназы фосфорилазы (см. рис. 12-31), в норме ассоциированные с этими киназами, на схеме не показаны.
|
|
|
Заключение
Известны три основных семейства поверхностных клеточных рецепторов, передающих внеклеточные сигналы различными способами.
Рецепторы, образующие канал, - это чувствительные к нейромедиатору ионные каналы, которые быстро открываются или закрываются в
ответ на связывание нейромедиатора и изменяют электрическую возбудимость клетки. Каталитические рецепторы являются в основном
тирозин-специфическими киназами, которые прямо фосфорилируют определенные белки
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
клеток-мишеней по остаткам тирозина. Рецепторы, сопряженные с G-белками, опосредованно активируют или инактивируют ферменты или
ионные каналы, связанные с плазматической мембраной, через регуляторные GTP-связывающие белки (G-белки). Эти G-белки «выключают» сами
себя, медленно гидролизу я связанный GTP. Некоторые из таких рецепторов активируют или ингибируют аденилатциклазу, изменяя этим
концентрацию внутриклеточного посредника сАМР. Другие активируют фосфоинозитид-специфичную фосфолипазу С, которая гидролизует фос-
фатидилинозитол-бисфосфат (Р1Р2), образуя из него два внутриклеточных посредника: 1) инозитолтрисфосфат (InsP3), освобождающий ионы
Са2+ из их внутриклеточных хранилищ и повышающий их концентрацию в цитозоле; и 2) диацилглицерол, остающийся в плазматической мембране
и активирующий протеинкиназу С, которая фосфорилирует различные клеточные белки. Разнообразные ответы, вызываемые активацией
рецепторов, сопряженных с G-белками, быстро прекращаются после удаления внеклеточного сигнального лиганда. При этом G-белки
|
|
|
инактивируются сами, InsP3 быстро дефосфорилируется фосфатазой, диацилглицерол превращается в фосфатидную кислоту или расщепляется
с образованием арахидоновой кислоты, сАМР гидролизуется фосфодиэстеразой, а ионы Са2 + быстро откачиваются из цитозоля.
12.4. Механизм действия циклического AMP и ионов кальция [22]
Так как сАМР и Са2+-возможно, самые важные внутриклеточные посредники, появляющиеся в результате активации рецепторов клеточной поверхности, механизм их действия заслуживает специального рассмотрения. Оба они действуют как аллостерические эффекторы -
активируют определенные белки, присоединяясь к ним и изменяя их конформацию.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 608; Нарушение авторских прав?; Мы поможем в написании вашей работы!