КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Кадгерины - семейство гомологичных гликопротеинов клеточной поверхности - осуществляют у
|
|
|
|
позвоночных Са2+-зависимую межклеточную адгезию [34]
Иммунологические методы, представленные на рис. 14-63, сыграли также решающую роль в открытии трех родственных гликопротеинов клеточной поверхности, называемых кадгеринами, которые участвуют в Са2+-зависимой межклеточной адгезии у позвоночных. Е-кадгерин имеется на поверхности многих эпителиальных клеток (и в клетках зародышей млекопитающих до имплантации), N-кадгеринка поверхности нервных клеток, клеток сердца и хрусталика, а Р-кадгерин — на. клетках плаценты и эпидермиса; как и N-CAM, все они иногда встречаются и в других тканях в процессе развития. Эти три кадгерина являются гомологичными трансмембранными гликопротеинами, лишь однократно проходящими через мембрану (каждый из них состоит примерно из 700 аминокислотных остатков), и в этом отношении они сходны с N-CAM. Однако в отсутствие Са2+ конформация кадгеринов сильно изменяется и поэтому они быстро расщепляются протеолитическими ферментами. Поскольку некоторые клетки, например эндотелиальные,
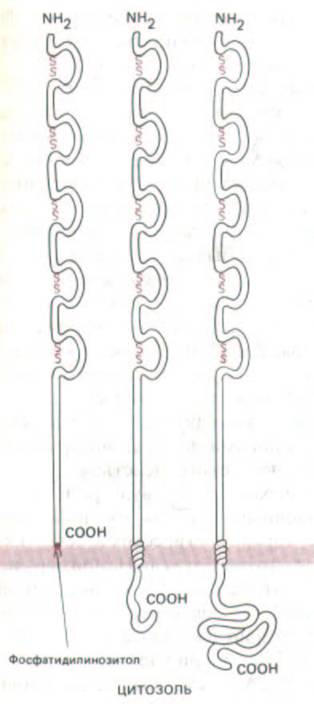
Рис. 14-66. Схематическое изображение трех форм N-CAM. Во всех трех случаях внеклеточная часть полипептидной цепи одинакова и организована в виде пяти доменов, сходных с доменами иммуноглобулинов. Каждый такой домен представляет собой петлю, концы которой связаны дисульфидными мостиками. (По данным В. A. Cunningham et al. Science 236: 799-800, 1987. Copyright 1987 by the AAAS.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
проявляют Са2 +-зависимую адгезию, но при этом не экспрессируют ни одного из трех известных кадгеринов, можно ожидать, что будут открыты новые представители семейства кадгеринов.
|
|
|
Лучше всех охарактеризован Е-кадгерин, называемый также молекулой адгезии печеночных клеток (L-CAM) или увоморулином. Большая внеклеточная часть его полипептидной цепи образует три гомологичных домена, по всей видимости не родственных доменам иммуноглобулинов.
Вероятно, он играет важную роль в скреплении клеток различных эпителиев. Например, Са2 +-зависимая реагрегация диссоциированных эпителиальных клеток печени блокируется антителами к Е-кадгерину. В зрелых эпителиальных тканях Е-кадгерин обычно концентрируется в адгезионных поясах, где он, как полагают, служит трансмембранным линкером, связывающим кортикальные актиновые цитоскелеты клеток, удерживая их вместе (разд. 14.1.3). Он также участвует в компактизации бластомеров в раннем зародыше мыши (разд. 16.2.4). Во время компактизации вначале рыхло расположенные бластомеры прижимаются друг к другу, плотно упаковываются и связываются межклеточными соединениями. Антитела к Е-кадгерину блокируют компактизацию бластомеров, тогда как антитела, реагирующие со многими другими поверхностными молекулами этих клеток, не оказывают такого действия.
Кажется вероятным, что кадгерины играют ключевую роль и на более поздних стадиях развития позвоночных, так как их появление и исчезновение коррелирует с важными морфогенетическими событиями, при которых ткани отграничиваются друг от друга. Например, по мере формирования нервной трубки и отделения ее от покровной эктодермы (разд. 16.1.9) в клетках развивающегося нервного эпителия исчезает Е-
кадгерин и появляется N-кадгерин (а также и N-CAM) (рис. 14-67). Когда клетки нервного гребня мигрируют из нервной трубки, они теряют N-
кадгерин (как и N-CAM, см. выше), но вновь начинают вырабатывать его позднее, при формировании нервного ганглия (см. рис. 14-56).
|
|
|
Биологическое значение поразительной зависимости белков межклеточной адгезии из семейства кадгеринов от концентрации ионов кальция не известно. Например, еще нет никаких данных о том, что для контроля межклеточной адгезии в процессе развития осуществляется регуляция внеклеточной концентрации кальция.
14.3.8. Молекулы клеточной поверхности, участвующие в адгезии между клетками и между клетками и матриксом, можно рассматривать как элементы морфогенетического кода [35]
Цитофизиологические, морфологические и биохимические исследования указывают на то, что клетка даже одного определенного типа использует много различных молекулярных механизмов прикрепления к другим клеткам и к внеклеточному матриксу. Некоторые из этих механизмов связаны со специализированными межклеточными соединениями, а другие - нет (рис. 14-68). Поскольку отдельная клетка использует большое число адгезивных систем, почти у каждого типа клеток найдется хотя бы одна система межклеточной адгезии, общая с любым другим типом, и поэтому все клетки будут обладать некоторым сродством Друг к другу. Обычно клетки разных тканей (и даже от весьма далеких видов)
способны образовывать друг с другом десмосомы, щелевые контакты и адгезионные соединения. Это позволяет предполагать, что участвующие в таких соединениях белки высококонсервативны (идет ли речь о разных тканях или видах). Однако точно так же, как каждая клетка многоклеточного животного содержит определенный набор поверхност-
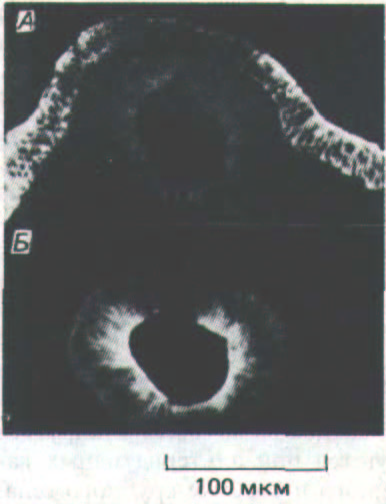
Рис. 14-67. Иммунофлуоресцентные микрофотографии поперечного среза куриного эмбриона: развивающаяся нервная трубка помечена антителами к Е-кадгерину (А) и к N-кадгерину (5). Обратите внимание, что клетки лежащей выше эктодермы содержат только Е-кадгерин, а клетки нервной трубки утратили Е-кадгерин, но в них появился N-кадгерин. (С любезного разрешения Kohei Hatta и Masatoshi Takeichi.)
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
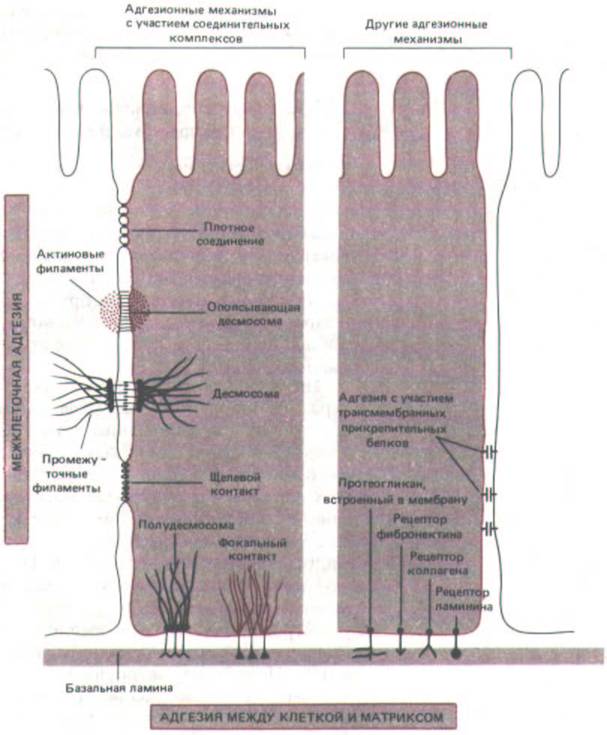
Рис. 14-68. Обобщенная схема адгезионных механизмов, используемых типичными эпителиальными клетками для прикрепления друг к другу и к внеклеточному матриксу (базальной мембране). Слева представлены механизмы с участием специализированных областей, видимых при электронной микроскопии обычных препаратов и (или) препаратов, полученных методом замораживания-скалывания. Справа иные механизмы. В некоторых случаях в соединении клеток между собой или с матриксом при помощи тех и других механизмов участвуют одни и те же гликопротеины клеточной поверхности. Как указывалось в тексте, все специализированные адгезионные механизмы, за исключением щелевых контактов, являются Са2 +-зависимыми; из остальных адгезионных механизмов лишь некоторые зависимы от Са 2+
|
|
|
ных рецепторов, дающий ей возможность специфическим образом реагировать на комплементарный набор растворимых сигнальных молекул (гормонов или локальных медиаторов), так и каждая клетка в ткани обладает определенной комбинацией (или концентрацией) рецепторов, позволяющих ей связываться определенным специфическим образом с другими клетками или с внеклеточным матриксом.
В отличие от рецепторов для растворимых веществ, которые связывают свои специфические лиганды с высоким сродством, рецепторы, связывающие молекулы клеточной поверхности или внеклеточного матрикса, осуществляют это с относительно низким сродством. Поэтому действие этих рецепторов основано на многократном увеличении силы связывания за счет одновременного соединения многих рецепторов со многими лигандами соседней клетки или внеклеточного матрикса. Поскольку у каждых двух клеток имеется некоторый спектр специфических рецепторов адгезии для других клеток и для матрикса, а также их концентраций и распределения по клеточной поверхности, то это и будет определять суммарное сродство, с которым клетки связываются друг с другом и с матриксом. Можно полагать, что именно этот спектр и есть тот «морфогенетический код», который определяет, как клетки будут организованы в ткани. Поскольку животные клетки даже близко родственных типов правильно рассортировываются in vitro, они должны быть способны определять относительно малые различия в адгезивных свойствах и использовать эти различия для установления лишь наиболее
|
|
|
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К. Уотсон Дж. Д. Молекулярная биология клетки: В 3-х т. 2-е изд. перераб. и доп. Т. 2.: Пер. с англ. – М.: Мир, 1993. – 539 с.
адгезивных из многих возможных контактов с другими клетками и матриксом. Наблюдения над подвижными клетками в культуре позволяют предполагать, как бы это могло осуществляться.
14.3.9. Высокоподвижные клетки служат чувствительными детекторами малых различий в адгезивности [36]
Клетки, участвующие в морфогенетических процессах у зародыша, часто очень подвижны. Если такие клетки диссоциировать и поместить в культуральную чашку, то вначале они будут по всем направлениям выпускать микрошипы и ламеллоподии, а затем активно расползаться по поверхности чашки. Эта подвижность часто совпадает с появлением различий между клетками и, следовательно, с периодом, когда важную роль должны будут играть процессы клеточного узнавания. Например, в зародыше Xenopus клетки внезапно становятся очень подвижными на стадии перехода к средней бластуле, когда начинается транскрипция генов (разд. 16.1.2).
Интенсивное изучение клеточной подвижности проводилось на культурах фибробластов, нейтрофилов и регенерирующих нейронов. Его результаты, суммированные в гл. И, указывают на то, что подвижные клетки являются чрезвычайно чувствительными детекторами малых различий в адгезивности. Микрошипы и ламеллоподии, выпускаемые во всех направлениях, по-видимому, участвуют в процессе «перетягивания каната», в результате которого клетка поляризуется и уверенно движется в направлении наиболее адгезивной части субстрата, даже если различия в адгезивности очень малы (разд. 11.6.3). Фибробласты, например, будут неуклонно двигаться вверх по малому градиенту адгезивности, создавшемуся на поверхности культуральной чашки. Изучение хемотаксиса у нейтрофилов позволяет предполагать, что подвижная клетка способна выявлять различия в адгезивности по обеим сторонам клетки всего лишь в 1%. Подобным же образом клетки в тканях могли бы с высокой чувствительностью расшифровывать «морфогенетический код» на клеточных поверхностях, уверенно двигаясь для установления тесного контакта с теми "из соседних клеток, к которым они наиболее адгезивны.
|
|
|
|
|
Дата добавления: 2015-08-31; Просмотров: 669; Нарушение авторских прав?; Мы поможем в написании вашей работы!