КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Регуляция численности популяций в биоценозах 2 страница
|
|
|
|
Для сибирского шелкопряда известен также эффект группы – положительное влияние оптимальной плотности, которое проявляется в основном у гусениц I–III возрастов и выражается в ускорении их развития более чем в 2 раза. В регуляцию хода численности вредителя вносит вклад также гетероцикличность – наличие особей с одно– и двухлетним циклом в зависимости от плотности популяции.
Повышение плотности очага сильно ускоряет метаболические процессы у членов популяции, что выражается как в ускорении сроков развития, так и в изменении поведения и внешнего вида бабочек и гусениц – появляются особи с темной окраской. В разных фазах цикла массового размножения количество темных бабочек в лесах Средней Сибири изменяется от 10 до 80 %.
Среди естественных врагов сибирского шелкопряда отмечено более 60 видов. Однако, несмотря на разнообразие, энтомофаги не могут постоянно держать под контролем численность вредителя из-за высокой инерционности их количественной реакции. Усложняющий фактор – высокая активность сверхпаразитов. Многие энтомофаги сибирского шелкопряда гораздо менее его устойчивы к факторам абиотической среды. Например, один из эффективных паразитов яйцеед-теленомус в массе гибнет в холодные зимы. Вот почему вспышки размножения сибирского шелкопряда часто возникают после холодных малоснежных зим.
При низкой численности сибирского шелкопряда в резервациях за счет естественных врагов-энтомофагов гибнет до 95 % яиц, 40 % гусениц и 70 % куколок. Основными регуляторами при этом являются яйцеед-теленомус и тахина-мазицера. Общие показатели смертности в популяциях достигают более 99 %. Это свидетельствует о высоких возможностях вредителя переходить к вспышке. Быстрый рост численности может быть обеспечен даже небольшим (5-10 %) увеличением выживаемости. На фазе нарастания численности поражаемость от энтомофагов падает (яйца – до 50 %, гусеницы – до 11 %, куколки – до 23 %). Энтомофаги «не успевают» сдерживать растущую популяцию вредителя. В период кульминации вспышки пресс энтомофагов на популяцию шелкопряда усиливается и достигает наибольшего выражения в период разреживания и депрессии численности, когда естественные враги полностью уничтожают избыток особей.
|
|
|
Таким образом, при массовых вспышках размножения лесные насекомые-вредители выходят из-под межвидового контроля и их возврат к стабильной численности происходит в основном в результате внутрипопуляционных регулирующих механизмов, которые наиболее эффективно действуют лишь при очень высоких уровнях плотности, губительной для лесного насаждения.
Понимание и учет всех этих факторов, определяющих динамику численности лесных вредителей, совершенно необходимы для принятия действенной системы мер борьбы с ними и профилактики вспышек, для определения необходимости, сроков, доз и форм применения средств, сдерживающих массовое размножение. С экологической точки зрения задача заключается в удержании популяции в зоне депрессии численности.
Понимание механизмов, определяющих движение численности популяций, дает в руки ключ управления ими. Однако эта задача не может решаться трафаретно для разных видов. Во всех случаях необходимо детальное знание экологических особенностей конкретного вида и условий его биотического окружения, а также характер взаимодействия модифицирующих факторов среды.
8.6.5. Влияние ослабления или усиления пресса хищников на динамику популяций и структуру сообществ
Ярким примером того, к чему приводит снятие пресса потребителей на популяции жертв, являются масштабные «нашествия» чужеродных видов, попадающих в подходящие для размножения условия на других материках. На родине такие виды испытывают влияние активных регуляторов, занимают соответственные экологические ниши в биоценозах и, как правило, не считаются вредителями в хозяйственной практике человека. В новой среде, не имея специализированных врагов, эти виды за короткие сроки дают взрывы численности, нарушая структуру сложившихся сообществ. С экологических позиций впервые обзор и анализ таких событий был сделан английским зоологом Ч. Элтоном, одним из основоположников популяционной экологии. Вводя понятие «взрыв» применительно к непомерному увеличению численности какого-либо вида, Ч. Элтон объяснял, что он намеренно употребляет это слово, так как оно обозначает «внезапное освобождение сил, проявление которых сдерживалось ранее другими силами».
|
|
|
«Нашествия» иноземных видов, занос которых преднамеренно или случайно осуществляется человеком, широко распространены в современном мире и часто вызывают большие осложнения в хозяйственной деятельности. Относительно немногие примеры успешной борьбы с ними связаны, как правило, с интродукцией естественных врагов, т. е. воссозданием системы биоценотической регуляции. Если на «захваченной» видом территории постепенно складывается новый комплекс регуляторов, тогда его численность идет на убыль. Однако для некоторых вселенцев этот процесс происходит достаточно медленно. Таким примером является опасный вредитель картофеля колорадский жук Leptinotarsa septemlineata, распространение которого в Европе началось после заноса из Америки в 20-х годах прошлого века с западных берегов Франции. Постепенно распространяясь на восток, колорадский жук занял в настоящее время всю европейскую территорию, картофелеводческие районы Сибири, обнаружен на Дальнем Востоке и острове Сахалин (рис. 141). Жук и его личинки имеют ядовитую гемолимфу, спасающую их от насекомоядных птиц и других позвоночных. У колорадского жука на евроазиатской территории до сих пор не сформировался эффективный комплекс потребителей. Потери урожая картофеля от колорадского жука составляют в среднем до 40 %.

Рис. 141. Распространение колорадского жука по территории России (на 1990 г.)
|
|
|
Другим примером современного «нашествия» вида, изменившего всю структуру сообщества, является размножение в Черном море гребневика Mnemiopsis leigyi, занесенного с балластными водами судов от побережья Северной Америки (рис. 142). Этот гребневик обладает гигантским репродуктивным потенциалом, производя ежесуточно до 10 000 яиц. Личинки тоже могут продуцировать яйца, из которых развиваются взрослые гребневики. При наличии пищи общая масса мнемиопсиса может удваиваться за 2–3 суток. Гребневик питается ветвистоусыми и веслоногими рачками, другим мелким зоопланктоном, икрой и личинками рыб. Впервые мнемиопсис был отмечен в Черном море в 1987 г. Уже через три года его масса составила 3–4 кг, а местами до 12 кг под каждым квадратным метром воды. Одновременно резко упала численность медуз Aurelia aurita, господствовавших до этого в планктоне (рис. 143). Выедая кормовой планктон, икру и личинок рыб, гребневик вызвал резкое падение общего их улова. Например, вылов ставриды сократился, со 110–115 тыс. т всего до 3 тыс. т. В Азовском море запасы тюльки – корма для хищных рыб – сократились в 400 раз, а ее добыча упала с 50–70 тыс. т до 100 т. В последующие годы численность гребневика понизилась, но он по-прежнему выедал основную массу зоопланктона (рис. 144). С 1997 г. в Черном море стал встречаться другой гребневик – Веrое ovata, распространенный на севере Атлантики и в Баренцевом море. Для него мнемиопсис явился хорошей кормовой базой, и численность последнего упала на порядок величин. В некоторых местах акватории он вообще не встречался. Снова стала размножаться медуза аурелия. Однако обилие Beroe в Черном море неустойчиво, этот гребневик адаптирован к более высокой солености воды. Появление и размножение его происходит волнами, а в период спада снова размножается Mnemiopsis.
Рис. 142. Гребневик мнемиопсис – внешний вид (из И. Е. Виноградова и Э. А. Шукшиной, 1993)

Рис. 143. Колебания биомассы аурелии и мнемиопсиса в глубоководной части Черного моря (в пересчете на углерод, по И. Е. Виноградову и Э. А. Шукшиной, 1993)
|
|
|

Рис. 144. Колебание биомассы в открытых районах Черного моря некоторых групп планктонных организмов, которыми питается мнемиопсис (по И. Е. Виноградову и Э. А. Шукшиной, 1993): 1 – саггиты; 2 – веслоногий рачок Calanus luxinus; 3 – мелкие рачки и личинки донных животных
Чрезмерное усиление пресса хищников на популяцию какого-либо вида в сообществе может отозваться через сеть трофических зависимостей на довольно далеких уровнях. Например, можно выстроить цепь причинных связей между усилением промысла минтая и других рыб в районе Берингова моря и исчезновением зарослей прибрежных водорослей – ламинарий. Усиленный рыбный промысел (при котором человек выступает в роли мощного хищника) стал одной из причин падения численности питающихся рыбой сивучей и нерпы – основной добычи крупных хищных дельфинов – косаток. С 1991 г. косатки стали охотиться в прибрежных водах на каланов, на которых не нападали раньше, добывая пищу в открытом океане. С начала 90-х годов на Алеутских островах исчезло 40 тыс. каланов, являющихся заповедным видом. Численность этих зверей за 7 лет сократилась почти в 10 раз. Каланы питаются в основном морскими ежами, обилие которых в результате возросло тоже на порядок величин. Морские ежи – главные враги бурых водорослей, заросли которых у побережья сократились за 10 лет в 12 раз.
8.6.6. Математическое моделирование в экологии
Надорганизменные системы, которые изучает экология – популяции, биоценозы, экосистемы, – чрезвычайно сложны. В них возникает множество взаимосвязей, сила и постоянство которых непрерывно меняются. Одни и те же внешние воздействия могут привести к различным, иногда прямо противоположным результатам, в зависимости от того, в каком состоянии находилась система в момент воздействия.
Предвидеть ответные реакции системы на действие конкретных факторов можно лишь через сложный анализ существующих в ней количественных взаимоотношений и закономерностей. В экологии поэтому широкое распространение получил метод математического моделирования как средство изучения и прогнозирования природных процессов.
Суть метода заключается в том, что с помощью математических символов строится абстрактное упрощенное подобие изучаемой системы. Затем, меняя значение отдельных параметров, исследуют, как поведет себя данная искусственная система, т. е. как изменится конечный результат.
Модели строят на основании сведений, накопленных в полевых наблюдениях и экспериментах. Чтобы построить математическую модель, которая была бы адекватной, т. е. правильно отражала реальные процессы, требуются существенные эмпирические знания. Отразить все бесконечное множество связей популяции или биоценоза в единой математической схеме нереально. Однако, руководствуясь пониманием, что в надорганизменных системах имеется внутренняя структура и, следовательно, действует принцип «не все связи существенны», можно выделить главные связи и получить более или менее верное приближение к действительности.
В построении математических моделей сложных процессов выделяются следующие этапы.
1. Прежде всего, те реальные явления, которые хотят смоделировать, должны быть тщательно изучены: выявлены главные компоненты и установлены законы, определяющие характер взаимодействия между ними. Если неясно, как связаны между собой реальные объекты, построение адекватной модели невозможно. На этом этапе должны быть сформулированы те вопросы, ответ на которые должна дать модель. Прежде чем строить математическую модель природного явления, надо иметь гипотезу о его течении.
2. Разрабатывается математическая теория, описывающая изучаемые процессы с необходимой детальностью. На ее основе строится модель в виде системы абстрактных взаимодействий. Установленные законы должны быть облечены в точную математическую форму. Конкретные модели могут быть представлены в аналитической форме (системой аналитических уравнений) или в виде логической схемы машинной программы. Модель природного явления есть строгое математическое выражение сформулированной гипотезы.
3. Проверка модели – расчет на основе модели и сличение результатов с действительностью. При этом проверяется правильность сформулированной гипотезы. При значительном расхождении сведений модель отвергают или совершенствуют. При согласованности результатов модели используют для прогноза, вводя в них различные исходные параметры.
Следует, однако, отметить, что сама по себе математическая модель не может служить абсолютным доказательством правильности той или иной гипотезы, так как может оказаться, что разные гипотезы приводят к сходным результатам, но она служит одним из путей анализа реальности.
Расчетные методы в случае правильно построенной модели помогают увидеть то, что трудно или невозможно проверить в эксперименте, позволяют воспроизводить такие процессы, наблюдение которых в природе потребовало бы много сил и больших промежутков времени. В математических моделях можно «проигрывать» разные варианты – вычленять разные связи, комбинировать отдельные факторы, упрощать или усложнять структуру систем, менять последовательность и силу воздействий – все это дает возможность лучше понять механизмы, действующие в природных условиях.
Моделируют различные по характеру процессы, происходящие в реальной среде, как, например, отдельные типы экологических взаимодействий хищник – жертва, паразит – хозяин, конкурентные отношения, мутуализм и др. Математическими моделями описываются и проверяются разные варианты динамики численности, популяций, продукционные процессы в экосистемах, условия стабилизации сообществ, ход восстановления систем при разных формах нарушений и многие другие явления. Сами методы математического моделирования биологических систем развиваются, совершенствуются и разнообразятся.
Например, одну из простейших математических моделей для системы паразит – хозяин в динамике численности насекомых разработал в 1925 г. статистик А. Лотка, который вывел следующие уравнения:
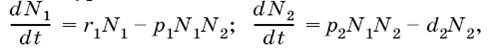
где N1 – численность популяции хозяина; N2 – численность популяции паразита; r1 – удельная скорость увеличения популяции хозяина; d2 – удельная скорость гибели популяции паразита; p1 и р2 – константы. График процесса паразитической инвазии, построенный по таким уравнениям, обнаруживает, что в результате взаимодействия двух видов должны возникать осцилляции (колебания) с постоянной амплитудой, которая зависит от соотношения между скоростями увеличения численности двух видов.
В это же время математик В. Вольтерра выявил сходные закономерности для системы хищник – жертва, обрабатывая статистические данные рыбного промысла. Один из выведенных им законов – «закон периодического цикла» – гласит, что процесс уничтожения одного вида другим может привести к периодическим колебаниям численности популяций обоих видов, зависящих только от коэффициентов роста популяций хищника и жертвы и от исходной относительной численности.
В период, когда были сделаны эти расчеты, экологи вели поиск причин циклических колебаний численности, которые были обнаружены к тому времени у ряда видов. Делались попытки отыскать внешние факторы (космические, солнечные, атмосферные), ответственные за периодические изменения популяций. Модели А. Лотки и В. Вольтерра позволили выдвинуть идею, что периодический колебательный режим в популяциях может возникнуть в результате межвидовых отношений и без внешнего периодического воздействия. Эта идея оказалась плодотворной для дальнейшего развития теории динамики численности популяций. Однако сама модель являлась не адекватной, т. е. не описывала действительность, так как в природе практически не обнаруживаются подобные непрерывные осцилляции с постоянной амплитудой у пар видов, связанных по типу хищник – жертва или паразит – хозяин.
Уравнения А. Лотки и В. Вольтерра были чрезвычайно упрощенными, так как исходили из целого ряда нереальных допущений: что изменение численности популяции одного вида немедленно вызывает ответную реакцию популяции другого вида, что «аппетиты» хищника беспредельны, поиски жертв случайны, что плодовитость хищников пропорциональна численности всей популяции жертв.
Как показал Г. Ф. Гаузе (1934, 1935), даже в условиях упрощенного эксперимента с простейшими трудно добиться соблюдения этих допущений. В его опытах с инфузориями удалось получить лишь два цикла хищник – жертва, после чего система пришла к разрушению. В природе колебания численностей имеют более сложный характер. Во взаимодействиях хищника и жертвы широко распространен эффект «запаздывания» из-за разницы в скоростях размножения, играют роль такие показатели, как степень насыщения («функциональная реакция») хищников, время, затрачиваемое ими на поиск и поимку добычи, способность переключаться на другую пищу, защитные приспособления жертв, размещение их в пространстве и территориальное поведение, возрастная и половая структура популяций и многое другое. Кроме того, рост численности популяций может сдерживаться и другими причинами, в том числе внутривидовыми взаимоотношениями.
В 1933 г. А. Никольсон, несколько усложнив математическую модель Лотки и введя в систему дополнительных хозяев и паразитов, показал, что это ослабляет осцилляции. В 1936 г. А. Н. Колмогоров разработал новые подходы и описал также возможности устойчивого стационарного состояния системы взаимодействующих через трофические связи видов. Позднее для систем хищник – жертва, паразит – хозяин было предложено множество других моделей. С введением в модели дополнительных параметров сильно усложняется математический аппарат и техника расчетов. Многие из этих ограничений позволило снять использование электронно-вычислительных машин.
В экологии сначала преобладали математические модели, основанные на предположениях о существовании в природе четких причинно-следственных зависимостей между популяциями в сообществах (так называемый детерминистский подход). В настоящее время меняется сам подход к математическому моделированию в экологии. Разработаны так называемые имитационные модели, основное внимание в которых уделяется именно разнообразию внутренней структуры популяций и сообществ. Вместо отбрасывания «несущественных» связей математики пытаются определить роль внутреннего разнообразия в поддержании существования надорганизменных систем.
Математическое моделирование широко применяется при решении экологических проблем, связанных с антропогенными воздействиями на природную среду. В современных математических моделях выделяют тактические и стратегические модели. Тактические модели экосистем и популяций служат для экологического прогнозирования их состояния, в том числе при разного рода экзогенных воздействиях. Стратегические модели строят в основном с исследовательскими целями, для вскрытия общих законов функционирования биологических систем, таких, как стабильность, разнообразие, устойчивость к воздействиям, способность возвращаться в исходное состояние. В задачи стратегических моделей входит изучение с помощью ЭВМ последствий разных стратегий управления экосистемами, чтобы иметь возможность выбрать оптимальную.
Модели, которые описывают взаимодействие общества и природы и в которых учитывают не только экологические, но и экономические, демографические и социальные показатели, называют эколого-экономическими моделями. Такие модели разрабатывают для долгосрочного прогнозирования экономического роста и общей оценки влияния человеческой деятельности на природную среду.
Глава 9. ЭКОСИСТЕМЫ
|
|
|
|
|
Дата добавления: 2017-01-14; Просмотров: 213; Нарушение авторских прав?; Мы поможем в написании вашей работы!