КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Аеробне окислення вуглеводів
|
|
|
|
Анаеробне та аеробне перетворення вуглеводів тісно пов’язані між собою. Це виявляється у тому, що обидва процеси проходять однаково до стадії утворення ПВК; в них беруть участь одні і ті самі ферменти та утворюються однакові проміжні продукти. Відмінності між цими перетвореннями вуглеводів починаються з перетворення ПВК. В аеробних умовах піруват окисляється до вуглекислого газу та води в аеробній стадій катаболізму, яка називається диханням. Спершу піруват окисляється до ацетил-КоА та вуглекислого газу. Це відбувається під впливом ферментів, об’єднаних структурно в піруватдегідрогеназний комплекс. Це мультиферментна система, що знаходиться в еукаріотичних клітинах в мітохондріях, в прокаріотичних - в цитоплазмі.
СН3–СО–СООН + НАД + КоА–SН ®СН3СО~S–КоА + НАД×Н2 + СО2
Ця реакція проходить в декілька стадій. В ході її відбувається окислювальне декарбоксилювання пірувату, в результаті якого карбоксильна група пірувату видаляється у вигляді молекули вуглекислого газу, а його ацетильна група включається до складу ацетил-СоА. Утворений НАДН2 передає водень в дихальний ланцюг. Утворений ацетил-СоА вступає в цикл лимонної кислоти (цикл трикарбонових кислот, цикл Кребса), який, на відміну від гліколізу, що включає лінійну послідовність ферментативних реакцій, працює в циклічному режимі.
Реакції цикл Кребсу відбуваються у мітохондріях. Цикл трикарбонових кислот був відкритий англійським біохіміком Гансом Кребсом. За це видатне відкриття Кребс отримав Нобелівську премію в 1953 р. разом з Ф. Ліпманом. Цикл Кребсу є центром, в якому сходяться практично всі метаболітичні шляхи, це кінцевий шлях окислення вуглеводів, жирних кислот, амінокислот. Реакція, що каталізується цитрат-синтазою, представляє собою конденсацію ацетил-КоА та ЩОК, утворюється лимонна кислота, відбувається розщеплення тіоефірного зв’язку та вивільнення коферменту А. Звільнений SH-КоА може тепер брати участь у окислювальному декарбоксилюванні нової молекули пірувату з утворенням нової молеку
 |
ли ацетил-КоА.
|
|
|
Схема циклу Кребса
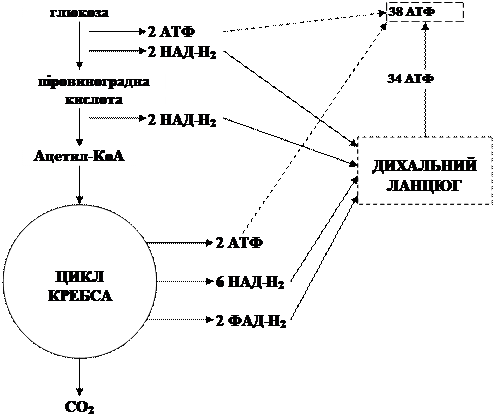
Фермент аконітаза каталізує оборотне перетворення лимонної кислоти в ізолимонну. Як проміжний продукт (в нормі він не відділяється від активного центру ферменту) утворюється трикарбонова цисаконітова кислота. В клітині ця реакція протікає зліва направо, оскільки продукт реакції (ізолимонна кислота) швидко захоплюється в наступні стадії циклу. Аконітаза містить залізо та кислотолабільні атоми сірки, згруповані в залізо-сірчистий центр. На наступній стадії циклу ізолимонна кислота дегідрується з утворенням a-кетоглутарової кислоти та вуглекислого газу під дією ізоцитратдегідрогенази. Існує два типи ізоцитратдегідрогенази: одна використовує як акцептор електронів НАД, а інша - НАДФ. Перший тип ферменту зустрічається лише в мітохондріях, другий в мітохондріях та в цитозолі. В циклі Кребса беруть участь обидва типи ферменту, але переважає НАД-залежна ізоцитратдегідрогеназа. Для її дії необхідні іони магнію або марганцю, а також специфічний активатор АДФ. Потім відбувається окислювальне декарбоксилювання a-кетоглутарової кислоти з утворенням високоенергетичної сполуки сукциніл-КоА та СО2, що каталізується a-кетоглутаратдегідрогеназним комплексом. Наступна реакція каталізується ферментом сукциніл-КоА-синтетазою. В ході цієї реакції сукциніл-КоА за участі ГДФ та неорганічного фосфату перетворюється в янтарну кислоту (сукцинат). Одночасно відбувається утворення високоергічного зв’язку ГТФ за рахунок високоергічного тіоефірного зв’язку сукциніл-КоА. Утворення ГТФ в даному випадку називається фосфорилювання на рівні субстрату, тому що джерелом необхідної енергії є окислення одного з органічних субстратів. ГТФ може потім передавати свою кінцеву фосфатну групу на АДФ з утворенням АТФ, ця оборотна реакція каталізується нуклеозид-дифосфат-кіназою. Потім янтарна кислота дегідрується з утворенням фумарової кислоти, що каталізується сукцинатдегідрогеназою, в молекулі якої з білком міцно (ковалентно) зв’язаний кофермент ФАД. Сам фермент міцно зв’язаний з внутрішньою мітохондріальною мембраною. Оборотна гідратація фумарату, внаслідок чого утворюється яблучна кислота (L-малат), каталізується фумарат-гідратазою (або ще відомий як фумараза). Фумараза високоспецифічна: вона гідратує лише транс-форму подвійного зв’язку фумарату і не діє на його цис-форму, а також ні на цис-, ні на транс-форму монокарбонових ненасичених кислот. В оборотній реакції фумараза проявляє специфічність у відношенні оптичних ізомерів; вона не здатна каталізувати дегідратацію D-малату. Кофермент для фумарази не потрібен. На останній стадій циклу лимонної кислоти НАД-залежна L-малатдегідрогеназа, що міститься в матриксі мітохондрій, каталізує дегідрування L-малату з утворенням ЩОК (оксалоацетату). Рівновага цієї реакції при стандартних умовах (тобто при концентраціях всіх компонентів 1 М та рН 7) сильно здвинута вліво. Але в клітині реакція іде зліва направо, тому що продукт реакції (оксалоацетат) швидко видаляється (витрачається в цитрат-синтазній реакції) і його реальна концентрація в клітині залишається весь час надзвичайно низькою. За один оборот циклу, що складається з восьми ферментативних реакцій, відбувається повне окислення (“згоряння”) однієї молекули ацетил-КоА. Для безперервної роботи циклу необхідне постійне поступання в систему ацетил-КоА, а коферменти (НАД і ФАД), що перейшли у відновлений стан, повинні знову окислятись. Окислення НАД . Н2 і ФАД . Н2 здійснюється в дихальному ланцюзі. Енергетика процесу: утворилося 4НАД×Н2 (з них 1НАДФН2), одній молекулі НАД×Н2 відповідає три молекули АТФ; 4´3=12молекул АТФ, 1ДАДН2=2 молекули АТФ, 1 молекула АТФ. Отже: 15 молекул АТФ´2 (бо 2 молекули ПВК)=30молекул АТФ.
|
|
|
Енергетичний ефект повного розщеплення глюкози:
|
|
|
Пентозний цикл (пентозофосфатний або гексозомонофосфатний шунт). Цей цикл забезпечує організм пентозами, які необхідні для синтезу нуклеїнових кислот тощо. У ссавців активність пентозного циклу відносно висока у печінці, серці, надниркових залозах, ембріональній тканині. Пентозний шлях активно реалізується в еритроцитах людини. Існує група спадкових хвороб людини, при яких активність деяких ферментів цього шляху знижена або взагалі відсутня. У таких хворих спостерігається гемоліз – руйнування еритроцитів з виділенням з них гемоглобіну, що веде до розвитку анемії. Стан різко погіршується під впливом деяких ліків (протималярійних препаратів). В Африці та Азії від цих хвороб страждають мільйони людей. Гексоза, перетворюючись в монофосфат, зразу окислюється в глюконову кислоту. Потім через ряд проміжних стадій утворюються фруктозомонофосфат та фосфогліцериновий альдегід. У подальшому ці продукти піддаються тим же перетворенням, що і при гліколітичному розпаді:
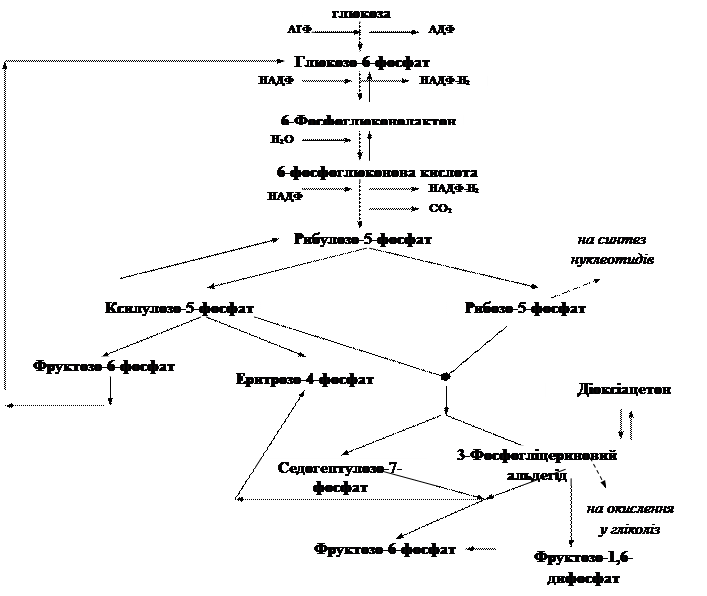 |
Основне призначення пентозофосфатного циклу:
· регенерувати у цитоплазмі відновлювач у формі НАДФ×Н2, що використовується при синтезі жирних кислот і стероїдів;
· в ньому синтезуються пентози, особливо D-рибоза, яка необхідна для синтезу нуклеїнових кислот;
· участь в утворенні глюкози з СО2 в темновій фазі фотосинтезу.
Важливе значення ферментів пентозофосфатного циклу полягає в тому, що вони разом з ферментами гліколізу створюють умови для взаємоперетворень 3-х, 4-х, 5-ти, 6-ти і 7-вуглецевих сахарів шляхом оборотного перенесення дво- або тривуглецевих фрагментів (глікоальдегідних або діоксіацетонових груп).
Отже, глюкозофосфатний шлях (пентозофосфатного циклу) є місцем перехрещення метаболічних шляхів, де утворюється значна кількість різних продуктів обміну речовин.
Ферментами фосфоглюконатного шляху може здійснюватися повне окислення глюкозо-6-фосфату до СО2 за таким рівнянням:
|
|
|
Глюкозо-6-фосфат + 12 НАДФ ® 6 СО2 + 12НАДФ×Н2 + Рн
Гліоксилатний цикл. У тварин він відсутній. Поширений у проростаючому насінні олійних культур. Таким шляхом із ацетильних груп, джерелом яких є жирні кислоти запасних триацилгліцеридів, утворюється глюкоза.Цей цикл є модифікацією циклу лимонної кислоти. Суть його полягає в тому, що ізолимонна кислота під впливом ізоцитрат-ліази розкладається на янтарну та гліоксилеву кислоти. Остання, взаємодіючи з ацетил-КоА під впливом малат-синтази, утворює яблучну кислоту. Яблучна кислота піддається подальшому окисленню, як у циклі Кребса. Жирним виділені реакції, які відрізняють цей цикл від цикла Кребса:
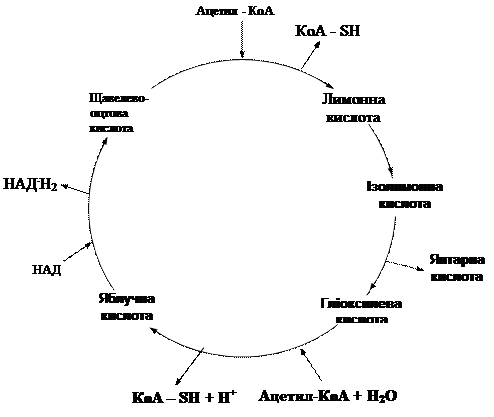
схема гліоксилатного циклу
Теорія біологічного окислювання
Згідно теорії біологічного окислення (теорії дихання), окислення починається з відняття водню піридиновими дегідрогеназами від субстрату (хН2) та передачі атомів водню флавіновим дегідрогеназам.
Наступним етапом є перенесення водню на убіхінон (коензим Q). Кожен водневий атом, що поступає від коферменту Q (убіхінону) розпадається на протон водню, який поступає в навколишнє середовище.та електрон, який поступає в систему цитохромів.
В цитохромах роль компоненту, що переносить електрони, відіграє залізо. Воно знаходиться у окисленій формі (Fе3+), але після приєднання електрону, переходить у відновлену форму (Fе2+).
Електрон від цитохрому b переходить до цитохрому с та далі до цитохромів а і а3 – міцному комплексу двох цитохромів, що називається цитохромоксидазою.
Будь-який цитохром може перенести лише по одному електрону, тому вважають, що у будь-якому дихальному ланцюзі є два ряди цитохромів.
Після цього система цитохромів передає електрони водню на кисень. В результаті приєднання електронів кисень переходить в іонну форму. Водень, що віддав електрони, також іонізується і переходить у розчин. Заряджені частки водню і кисню з'єднуються між собою, утворюючи воду. Оскільки при такому окисленні постійно має місце поглинання кисню, то його називають тканинним диханням.
Спрощена схема перенесення Н2 і електронів при окислювальному фосфорилюванні (дихальний ланцюг):
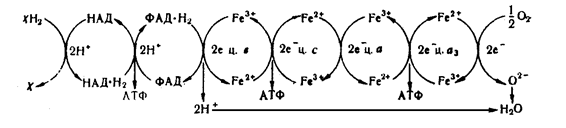 |
хН2 – субстрат, що окисляється
Біологічне значення перенесення водню й електронів полягає у використанні енергії утворення води. Реакція з'єднання водню і кисню – найбільш ефективна екзотермічна реакція між хімічними елементами. Природа використовувала цю реакцію як джерело енергії для біохімічних процесів. Поза організмом ця реакція відбувається миттєво, з вибухом, а в живому організмі – поступово. Водень передається кисню через ланцюг передавачів, і енергія виділяється не відразу, а по частинам, що дає можливість використовувати її для біохімічних процесів. Кожен етап переміщення водню та електронів від ферменту до ферменту супроводжується виділенням енергії і на певних етапах виділеної енергії достатньо для здійснення процесу фосфорилювання та утворення молекули АТФ. В усьому ланцюзі є три таких етапи. Процес утворення АТФ при біологічному окислюванні називається окислювальним фосфорилюванням. Цей шлях утворення АТФ є універсальним і використовується всіма організмами в процесі їхньої життєдіяльності в присутності кисню.
При перенесенні водню по дихальному ланцюгу від НАД . Н2 утворюється 3 молекули АТФ, при перенесенні від ФАД . Н2 – 2 АТФ.
Всі ферменти тканинного дихання пов’язані, головним чином, з мітохондріями. Нікотинамідаденіндинуклеотидні коферменти знаходяться в матриксі мітохондрій, металофлавопротеїни, убіхінон, цитохроми пов’язані з ліпідними структурами внутрішньої мембрани.
Запитання і вправи для самоконтролю.
1. Яка роль вуглеводів в організмі?
2. Зазначте основні шляхи розщеплення вуглеводів в організмі.
3. Охарактеризуйте основні типи бродіння і їх значення в харчовій промисловості.
4. Що таке гліколіз?
5. Назвіть реакції анаеробного перетворення вуглеводів.
6. Розрахуйте енергетичний баланс анаеробного розкладу вуглеводів.
7. Зазначте шляхи перетворення піровиноградної кислоти.
8. Опишіть хімізм спиртового бродіння.
9. Напишіть рівняння реакцій цикла Кребса.
10. Обчисліть загальний енергетичний ефект перетворення глюкози у СО2 і Н2О.
11. Охарактеризуйте гліоксилатний шлях обміну глюкози.
12. Яке значення пентозофосфатного катаболізму глюкози?
13. Що таке біолоігчне окислення? Охарактеризуйте тканинне дихання.
14. Як побудований дихальний ланцюг мітохонрій?
Список рекомендованої літератури: [1, с. 242-253, 320 – 352; 2, с. 197 – 224; 3, с. 196 – 217].
ОБМІН ЛІПІДІВ
|
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 6508; Нарушение авторских прав?; Мы поможем в написании вашей работы!