КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Физиологические и молекулярные механизмы адаптации к
|
|
|
|
Таким образом, с одной стороны, происходит торможение обменных процессов, как приспособление к выживанию в условиях низкотемпературного стресса, а с другой – идет такая перестройка дыхательного обмена, которая способствует накоплению метаболитов, участвующих в повышении зимостойкости. Переход на анаэробный тип дыхания наблюдается не только при низкой температуре, но и при кислородной недостаточности, засухе.
Второй этап – естественное закаливание т.е. возникновение физиологических и биохимических приспособлений в ходе естественного закаливания в осенний период. По теории Туманова закаливание происходит на свету в 2 фазы: при низких положительных температурах, а затем медленное охлаждение в условиях отрицательных температур. Закаливание направлено не предотвращение образования льда внутри клетки и повышение устойчивости к внутриклеточному льду (если он все-таки образовался) для уменьшения обезвоживания и механической деформации протопласта.
Таким образом, второй этап подготовки к зиме – это первая фаза периода закаливания. Она начинается при температурах +5-0°С. Озимые злаки проходят ее за 6-20 дней, древесные – за 30 дней. Подготовительные процессы закаливания не должны осуществляться быстро. Необходимо длительное, постепенное охлаждение, поскольку растение лучше успевает приспособиться к изменению температурного режима.
Что происходит в первую фазу закаливания при понижении температуры до 0°С? Необходимо, чтобы свободная вода из клетки быстро транспортировалась в межклетники. Это обеспечивается поддержанием высокой проницаемости мембран. Увеличению проницаемости, потере воды способствует трансформация белков и мембранных белков-ферментов. Однако определяющими становятся особенности липидного состава мембран растений. Изменяются физические и химические свойства мембран в направлении возрастания содержания ненасыщенных жирных кислот. Чем устойчивее растение, тем выше индекс ненасыщенности двойных связей жирных кислот липидов. При закаливании повышается содержание липидов, мембранных фосфолипидов и увеличивается их ненасыщенность. Накапливаются криопротекторы. Снижается точка замерзания цитоплазмы.
|
|
|
Увеличение содержания ненасыщенных жирных кислот обусловливает снижение температурного фазового перехода липидов из жидкостнокристаллического состояния в гель. У морозостойких растений она лежит ниже точки замерзания, а у неустойчивых она выше 0°С. Благодаря фазовым переходам мембран из жидкостно-кристаллического в твердое состояние снижается проницаемость мембран. Поэтому понижение температуры фазового перехода липидов у морозоустойчивых растений способствует сохранению высокой проницаемости мембран при
замораживании.
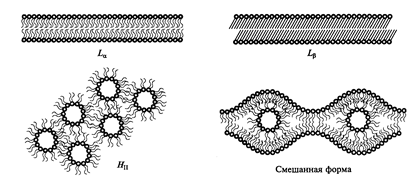
Рис. 1. Фазовые переходы липидов мембран при изменениях температуры и содержания воды в клетках: Lα – ламеллярная форма в жидкокристаллическом состоянии; Lβ – ламеллярная форма в твердом, гелеобразном состоянии; Н11 – гексагоналная форма; Н111 – смешанная форма
При понижении температуры происходит активация фосфолипазы А2, что приводит к накоплению свободных жирных кислот и вызывает изменения в энергетике клетки. Жирные кислоты в митохондриях становятся не только основными субстратами окисления, но и важнейшим регулятором – разобщителем дыхания и фосфорилирования, упрощающим превращение энергии субстратов в тепло (рис 2).
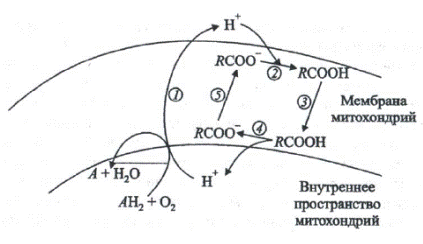
Рис. 2. Роль свободных жирных кислот в генерации тепла в
митохондриях при низкой температуре (по Скулачеву, 1998).
|
|
|
RCOOH, RCOO- – жирная кислота и ее анион, АН2 – дыхательный субстрат, А – продукт дыхания.
В обычных условиях откачивание H+ из внутреннего объема митохондрий наружу посредством ферментов ЭТЦ сопряжено с окислением дыхательных субстратов в продукты (1). При пониженной температуре откачанные из митохондрий ионы водорода на внешней поверхности мембраны присоединяются к анионам жирных кислот RCOO- (2).
Получающаяся при этом протонированная форма жирной кислоты
диффундирует через мембрану (3) и диссоциирует на ее внутренней
поверхности, давая RCOO- и Н+ (4). Протоны компенсируют внутри
митохондрий нехватку ионов водорода, удаляемых оттуда дыхательной цепью. Образующийся анион RCOO- возвращается наружу (5) при участии митохондриальных белков – анионных переносчиков, в частности АТФ/АДФ- антипортера. Его главная функция состоит в обмене аниона внешнего (цитозольного) АДФ на внутримитохондриальный анион АТФ, образованный Н+-синтазой. Антипортер хорошо различает гидрофильные анионы, например АДФ и ГДФ. Однако в отношении гидрофобных анионов такой избирательности не наблюдается. Поэтому АТФ/АДФ-антипортер оказывается способным к транспорту анионов жирных кислот. Разобщающий белок митохондрий, близкий к АТФ/АДФ-антипортеру, в отличие от последнего неспособен транспортировать нуклеотиды, но сохранил способность переносить анионы жирных кислот. При адаптации к низкой температуре разобщающий белок включается наряду с АТФ/АДФ-антипортером в транспорт анионов жирных кислот через митохондриальную мембрану. В результате конкурентного блокирования АТФ/АДФ-антипортера жирными кислотами транспорт АДФ и АТФ через внутреннюю мембрану митохондрий ингибируется, повышается активность альтернативного пути транспорта электронов и усиливается рассеивание энергии в виде тепла. Поэтому в тканях растений зимостойкой озимой пшеницы митохондрии генерируют тепло. Температура побегов пшеницы в течение некоторого времени остается более высокой, чем температура окружающей среды. Альтернативный путь в этот период выполняет защитную функцию, способствующую ускорению окисления.
В ходе закаливания происходит более масштабная перестройка дыхания: снижаются интенсивность поглощения кислорода и дыхательный коэффициент, увеличивается активность дегидрогеназ, что сопровождается появлением новых изоферментов (пероксидаз, каталаз, полифенолоксидаз, цитохромоксидаз). Понижение интенсивности дыхания происходит в меньшей степени у морозоустойчивых сортов. Повышение активности дегидрогеназ связано с усилением их роли в поставке НАДН, необходимых для усиливающихся в это время синтетических процессов, обмена и накопления защитных веществ (сахаров, аминокислот, белков).
|
|
|
Закаливание ведет также к появлению необычных форм митохондрий (гантелевидных, чашевидных, вплоть до образования митохондриальног ретикулума), с которыми связывают высокий уровень энергетического обмена.
У растений с ретикулярным типом организации митохондрий отмечена эффективная метаболическая адаптация, направленная на синтез криозащитных веществ.
отрицательным температурам:
Накопление криопротекторов – сахаров и других совместимых осмолитов. Известно, что при низкой температуре крахмал превращается в сахарозу, при потеплении происходит обратное превращение. Это превращение идет в клетках коры зимующих деревьев, в клубнях и корневищах. Сахара накапливаются в клеточном соке, органеллах, особенно в хлоропластах. Чем выше концентрация раствора, тем ниже точка его замерзания, поэтому при накоплении сахаров уменьшается количество образующегося льда. Сахара также препятствуют образованию или уменьшают количество токсических веществ, образующихся при обезвоживании клеток из-за образования льда.
Сахара являются главным дыхательным субстратом, являющимся источником энергии для синтезов, происходящих в первый период закаливания. Зимой сахара составляют 50-60% сухой массы в цитоплазме и органеллах. К полимерам-криопротекторам относятся также молекулы гемицеллюлоз (ксиланы, арабиноксиланы). Накапливаясь в клеточной стенке, они обволакивают кристаллы льда и тормозят их рост.
К криопротекторам относятся и многоатомные спирты: глицерол, сорбитол, маннитол; аминокислоты, например, пролин. Образование этих соединений приводит к снижению точки замерзания цитоплазмы.
|
|
|
Изменение состава мембранных липидов и увеличение текучести
мембран.
Защитной реакцией является увеличение количества ненасыщенных
жирных кислот в липидах, т.е. происходит активация десатураз.
Ограничение роста внеклеточного льда и синтез антифризных белков.
Растения могут синтезировать антифризные белки, которые тормозят рост кристаллов льда, не влияя при этом на температуру замерзания раствора.
Низкие температуры вызывают синтез антифризных белков, которые связываются с кристаллами льда, предотвращая или замедляя их дальнейший рост. Эти белки аккумулируются прежде всего в апопласте во время адаптации к холоду. Например, у растений озимой ржи обнаружено 6 антифризных белков с молекулярной массой от 16 до 35 кД. У чувствительных к морозу растениях такие белки обычно не обнаруживаются.
Синтез стрессовых белков холодового ответа.
В настоящее время идентифицировано несколько генов и кодируемых ими белков холодового ответа, которые часто обозначают как COR-белки.
Наиболее изучены COR-белки арабидопсиса, кодируемые четырьмя
семействами генов. Промоторы всех этих генов имеют одинаковый
регуляторный элемент CRT/DRE, с которым связывается транс-фактор СBF1.
Этот регуляторный белок может запускать экспрессию сразу всех COR-генов. Кодируемые этими генами белки крайне гидрофильны и сохраняют свою стабильность даже при кипячении. Один из COR-генов арабидопсиса экспрессируется в ответ на низкую температуру, засуху и АБК. Кодируемый этим геном белок синтезируется в цитоплазме и затем транспортируется в хлоропласт. Этот белок повышает криостабильность плазмалеммы, а также хлоропластных мембран, но не влияет на морозоустойчивость целого растения.
Повышение устойчивости к низким температурам всего растительного
организма зависит от одновременной работы всех имеющихся СOR-генов.
При адаптации к морозу синтезируются также аналоги LEA-белков,
которые важны также при адаптации растений к водному дефициту. Помимо этого синтезируются молекулярные шапероны, являющиеся гомологами БТШ- 90 и БТШ-70 и предотвращающие денатурацию белков при действии отрицательных температур.
Среди белков холодового стресса есть и ферменты антиоксидантной
защиты, различные гидролазы, а также ферменты биосинтеза веществ-
криопротекторов.
Во время второй фазы закаливания, когда происходит постепенное понижение температуры до –10-20°С и ниже со скоростью 2-3°С в сутки, в межклетниках образуется лед и начинают функционировать механизмы защиты от обезвоживания, подготовленные в течение первой фазы.
Осуществляются 2 основных приспособления. Во-первых, транспорт оставшейся после прохождения первой фазы закаливания свободной воды из клетки. Основную роль в транспорте свободной воды из клеток к местам внеклеточного образования льда играют мембраны. Высокая проницаемость мембран при замораживании обеспечивается увеличением содержания ненасыщенных жирных кислот. Во-вторых, повышение устойчивости клетки к обезвоживанию. Водоудерживающие силы увеличиваются за счет криопротекторов. Закаленные клетки не повреждаются концентрированным клеточным соком, т.к. в этот период они находятся в химически инертном состоянии. По мере усиления морозов объем клеток вследствие оттока воды уменьшается, а потом остается без изменений, несмотря на продолжение образования льда в межклетниках. Возможно, в этот период происходит замещение воды клеток воздухом, которое имеет большое значение для выживания растений, т.к. предохраняет клетки от деформации.
Когда наступает оттепель, растения утрачивают свойства, приобретенные
во время закаливания, и уровень их морозостойкости резко падает, причем происходит это всего за 1-2 дня. При последующем понижении температуры растения опять проходят вторую фазу закаливания и уровень их морозостойкости снова повышается. Однако если температура понизится очень быстро, растение может погибнуть. Поэтому перенесение растением зимнего периода определяется не только способностью выживать в условиях очень низких температур, но и зимостойкостью в целом, т.е. возможностью адаптироваться к повторяющимся оттепелям и заморозкам.
Таким образом, морозоустойчивость растений – состояние не постоянное, оно может изменяться в течение небольшого периода. Ее можно увеличить закаливанием, но можно и снизить действием определенных факторов (вода, температура, свет). Не все растения способны закаливаться, вследствие этого различные сорта озимых культур обладают неодинаковой морозоустойчивостью.
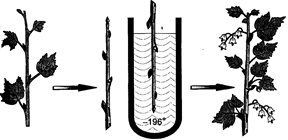 |
Древесные растения, развивающие морозоустойчивость в состоянии покоя, проходят
Рис.3. Закаленный побег смородины может плодоносить после пребывания при температуре -196оС (Туманову, 1979)
более сложный и длительный период закаливания. Подготовка к перезимовке у них начинается после окончания периода роста, задолго до наступления холодов. В течение второй половины лета в живых тканях накапливаются питательные вещества, главным образом крахмал, из которого осенью в результате гидролиза образуются моносахариды (невысокая устойчивость), или олигосахариды (более высокая устойчивость), или жирные масла (самая высокая). Эти вещества связываются с белками, образуя белково-липидный комплекс, который пропитывает цитоплазму. При понижении температур до области критических значений в клетках древесных растений происходит набухание цитоплазмы. Это состояние обратимо, и после оттаивания исходная структура восстанавливается. В это же время снижается содержание воды в тканях, происходит отток различных веществ из надземных органов в корневые системы (продукты обмена, гормоны роста). Клетки корней в свою очередь вырабатывают вещества, повышающие устойчивость организма против морозов. Благоприятным физиологическим условием для успешного закаливания является раннее прекращение роста, вызревание побегов и переход их в состояние покоя. Этому благоприятствуют короткий день, пониженная влажность и умеренное азотное питание.
|
|
|
|
|
Дата добавления: 2013-12-13; Просмотров: 663; Нарушение авторских прав?; Мы поможем в написании вашей работы!