КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Тема № 7. Инфекции и иммунитет. Пищевые заболевания (пищевые инфекции и пищевые отравления)
|
|
|
|
Тема №6 Важнейшие биохимические процессы превращений, вызываемые микроорганизмами: брожения - спиртовое, молочнокислое, масляно-кислое и их модификации; окислительные брожения - уксусно-кислое, лимонно-кислое; превращение белков- гниение.
Луи Пастером установлено, что изменения происходящие при брожении являются результатом жизнедеятельности микроорганизмов. В зависимости от преобладающих конечных продуктов различают типы брожений: спиртовое, молочно-кислое, масляно-кислое, их модификации и др. виды брожения.
Спиртовое брожение. Спиртовым брожением называют процесс превращения сахаров под действием ферментативной активности некоторых видов бактерий, мицеллиальных и дрожжевых грибов с накоплением в качестве основного продукта этилового спирта и углекислого газа:
С6Н12О6®2С2Н5ОН + 2СО2+ Q.
Но основными возбудителями этого вида брожения являются дрожжи (сахаромицеты). В анаэробных условиях превращение сахара в спирт происходит не сразу, процесс идет через ряд промежуточных реакций и протекает как бы в две стадии: первая (окислительная) включает превращение глюкозы до пировиноградной кислоты с образованием двух молекул восстановленного НАД×Н2 (первичный акцептор водорода):
С6Н12О6®2СН3СОСООН+2 НАД×Н2.
Под действием фермента пируватдекарбоксилазы дрожжи катализируют реакцию декарбоксилирования пировиноградной кислоты с отщеплением СО2 и образованием уксусного альдегида:
2СН3СОСООН®2СН3СНО+ 2СО2,
а вторая стадии (восстановительная) НАД×Н2 передает Н конечному акцептору (уксусный альдегид), который превращается в этиловый спирт:
2СН3СНО+ 2НАД×Н2 ®2С2Н5ОН+ НАД
С энергетической точки зрения брожение процесс мало экономичный, так как при сбраживании грамм-молекулы глюкозы синтезируется всего два моля АТФ.
|
|
|
Наряду с главными продуктами брожения в небольшом количестве образуются побочные продукты: глицерин, уксусный альдегид, уксусная и янтарная кислоты, сивушные масла (смесь высших спиртов) и некоторые другие вещества.
Общие условия спиртового брожения. Наиболее благоприятная концентрация сахара в среде для большинства дрожжей от 10 до 15%. При повышении концентрации сахара энергия брожения снижается, а при 30-35% брожение обычно прекращается, хотя в природе встречаются дрожжи, способные вызывать медленное брожение сахара даже при концентрации 60% и выше. Этиловый спирт, накапливающися в процессе брожения, неблагоприятно влияет на дрожжи. Его угнетающее действие проявляется уже при концентрации 2-5% в зависимости от вида и расы дрожжей. В большинстве случаев брожение прекращается при 12-14% (объемных) спирта. Получены расы, индуцирующие до 20% спирта.
Нормальное брожение протекает в кислой среде при рН 4-5. В щелочной среде при подщелачивании среды до рН 8 или при введении в среду бисульфита натрия направление брожения изменяется в сторону увеличения выхода глицерина (глицериновое брожение):
2С6Н12О6®2СН2ОНСНОНСН2ОН +С2Н5ОН +СН3СООН + 2СО2+ Q
По характеру брожения дрожжи подразделяют на верховые и низовые.
Брожение, вызываемое верховыми дрожжами, протекает бурно и быстро при температуре 20-28˚С. На поверхности бродящей жидкости образуется много пены, и под действием выделяющегося углекислого газа дрожжи выносятся в верхние слои субстрата. По окончании брожения дрожжи оседают на дно рыхлым слоем.
Брожение, вызываемое низовыми дрожжами, протекает спокойнее и медленнее, особенно при сравнительно низких температурах (4-10˚С). Газ выделяется постепенно, пены образуется меньше, дрожжи не выносятся на поверхность среды и быстро оседают на дно бродильной емкости.
|
|
|
При интенсивной аэрации среды дрожжевые грибы меняют тип энергетического обмена и переходят с процесса брожения на процесс дыхания, что называют эффектом Пастера: С6Н12О6 + 6О2 ®6СО2 +6Н2О + Q
Дыхание является более выгодным энергетическим процессом так как так как при сбраживании грамм-молекулы глюкозы синтезируется 36 молекул АТФ. Эффект Пастера используют при получении биомассы дрожжевых клеток для виноделия (ЧКД), хлебопечения (пекарские), в качестве кормового белка.
Процесс спиртового брожения лежит в основе хлебопечения, бродильных производств (виноделие, пивоварение, производство этилового спирта), глицерина. Совместно с молочно-кислым брожением используется при получении некоторых молочно-кислых продуктов (кефира, кумыса), при квашении овощей.
Молочно-кислое брожение. Это процесс превращения сахара в молочную кислоту по характеру брожения различают две группы молочно-кислых бактерий: гомоферментативные (типичные) и гетероферментативные (нетипичные).
Гомоферментативные бактерии образуют в основном (не менее 85-90%) молочную кислоту и очень мало побочных продуктов:
2С6Н12О6®2СН3СНОНСООН + Q
Гетероферментативные бактерии наряду с молочной кислотой образуют значительное количество других веществ:
2С6Н12О6®2СН3СНОНСООН +СООН (СН2)2 СООН+ С2Н5ОН + СН3СООН+СО2+ Q
Есть такие гетероферментативные молочно-кислые бактерии, которые кроме того продуцируют четырехуглеродные соединения ацетоин (СН3СНОНСОСН3) и диацетил (СН3СОСОСН3), обладающие своебразным приятным ароматом (специфическим для кисломолочных продуктов).
В зависимости от условий развития (рН, температуры, степени аэробности и др.) характер конечных продуктов брожения может меняться у одного и того же вида молочно-кислых бактерий. Процесс превращения глюкозы до пировиноградной кислоты у гомоферментативных молочно-кислых бактерий протекает по гликолитическому пути. Затем, в виду отсутствия фермента пируватдекарбоксилазы у этих бактерий, пировиноградная кислота не подвергается расщеплению и является конечным акцептором водорода под действием фермента лактатдегидрогеназы (лактикодегидрогеназы):
СН3СОСООН+ НАД×Н2. ®2СН3СНОНСООН + НАД
|
|
|
Превращение глюкозы гетероферментативными молочно-кислыми бактериями происходит по-иному, так как они отличаются набором ферментов. Из-за отсутствия у них фермента альдолазы изменяется начальный путь превращения глюкозы и процесс протекает не по гликолитическому пути, а по пентозофосфатному. После фосфорилирования глюкоза (глюкоза-6-фосфат) окисляется (отщепляется водород) и декарбоксилируется (отщепляется СО2). Образуется пентозофосфат, который фосфокетолазой расщепляется на фосфоглицериновый альдегид и ацетилфосфат. Фосфоглицериновый альдегид, как и у гомоферментативных молочнокислых бактерий, превращается в пировиноградную кислоту (ПВК), которая затем восстанавливается в молочную. Ацетилфосфат, в свою очередь, дефосфорилируется и через уксусный альдегид может превращаться: (алкогольдегидрогеназой) в этиловый спирт; (присоединяя О2) уксусную кислоту; ароматические соединения-(присоединяя СО2 и СН3СООН) ацетоин (СН3СНОНСОСН3) и из него (отщепляя 2Н2) в диацетил (СН3СОСОСН3)
Молочно-кислые бактерии имеют форму кокков (молочнокислые стрептококки) и палочек (лактобактерии), неподвижны, не образуют спор, грамположительны, являются факультативными анаэробами, некоторые микроаэрофилами. Различные молочнокислые бактерии образуют неодинаковое количество молочной кислоты. Преобладающее большинство гомоферментативных палочек продуцирует кислоты больше (до2-3.5%), чем стрептококки (ок. 1%). Поэтому палочковидные формы могут развиваться при рН 4,0 -3,8; кокковые формы при такой кислотности не развиваются. Наилучшая активность палочковидных бактерий проявляется при рН 5,5-6,0. Молочнокислые бактерии легко переносят высушивание, устойчивы к СО2 и этиловому спирту; многие виды существуют при содержании в среде до 10-15% этилового спирта.
По отношению к температуре молочнокислые бактерии делят на мезофилы (25-35˚С) и термофилы (40-45˚С), есть формы сохраняющиеся при нагревании до 85˚С в течение нескольких минут.
Наиболее важными в технологическом отношении являются:
|
|
|
- гомоферментативные мезофильные: Молочнокислый стрептококк (Lactococcus lactis) - образует антибиотик низин; сливочный стрептококк (Lactococcus cremoris) - образует антибиотик диплококцин;
- гомоферментативные термофильные: термофильный стрептококк (Streptococcus thermophilus); болгарская палочка (Lactobacterium bulgaricum);
-гетероферментативные: ароматобразующий стрептококк (Leuconostoc cremoris); кортокая палочка(Lactobacterium brevis); бифидобактерия (Bifidobacterium bifidum.
Молочно-кислые бактерии широко применятся в различных отраслях пищевой промышленности (особенно молочно-кислой). Большое значение эти бактерии имеют при квашении овощей, силосовании кормов, в хлебопечении при приготовлении ржаного хлеба, при производстве некоторых колбас и солено-вареных мясных изделий, при созревании слабо соленой рыбы, при получении молочной кислоты.
Спонтанно (самопроизвольно) возникающее молочно-кислое брожение в пищевых продуктах приводит к их порче: прокисание, помутнение, ослизнение.
Пропионово-кислое брожение. Это превращение сахара или молочной кислоты и ее солей под действием ферментативной активности пропионово-кислых бактерий в пропионовую и уксусную кислоты с выделением углекислого газа и воды:
3С6Н12О6. ®4СН3СН2СООН+ 2СН3СООН+ 2СО2 + 2Н2О+Q;
3 СН3СНОНСООН®2СН3СН2СООН+ СН3СООН+ СО2 + Н2О+Q
Некоторые пропионово-кислые бактерии могут образовывать, кроме того, муравьиную, янтарную и изовалериановую кислоты. При пропионово-кислом брожении превращение глюкозы до пировиноградной кислоты протекает также по гликолитическому пути. В дальнейшем пировиноградная кислот, претерпевая ряд превращений, восстанавливается в пропионовую. Пропионово-кислые бактерии относятся к актиномицетам. Это неподвижные, бесспоровые, грамположительные палочки слегка изогнутые, факультативные анаэробы.
Пропионово-кислое брожение является одним из важных процессов при созревании сычужных сыров.
Пропионовая кислота и ее соли служат ингибиторами мицеллиальных грибов и могут быть использованы для предотвращения плесневения пищевых продуктов. Некоторые виды пропионово-кислых бактерий применяют для получения витамина В12 (Propionibacterium freudenreichii subsp. shermanii).
Слизевое (декстрановое) брожение. Вызывают слизеобразующие бактерии, которые по своим свойствам близки к молочнокислым и способны образовывать вокруг клеточной стенки капсулу, состоящую из слизистых веществ (полисахариды, полипептиды). Процесс идет в несколько этапов:
1. ГИДРОЛИЗ САХАРОЗЫ:
С12Н22О11+ Н2О + ®С6Н12О6. +С6Н12О6.
сахароза глюкоза фруктоза
2. ПРЕВРАЩЕНИЕ ГЛЮКОЗЫ:
С6Н12О6. ®(С6Н10 О5)n+Н2О
глюкоза декстран
3. ПРЕВРАЩЕНИЕ ФРУКТОЗЫ
С6Н12О6®2СН3СНОНСООН +СООН 2(СН2)2 СООН+ С2Н5ОН +
фруктоза
+СН3СООН+ Q
Сбраживание глюкозы протекает по гетероферментативному молочно-кислому брожению.
Слизеобразующие бактерии вызывают ослизнение овощей, фруктов, хлебобулочных изделий (картофельная болезнь), сахарных сиропов, различных слабоалкогольных напитков и вина. Промышленного значения не имеет.
Маслянокислое брожение. Этот процесс представляет собой превращение сахара маслянокислыми бактериями в анаэробных условиях с образованием масляной кислоты, газа и водорода:
С6Н12О6. ®СН3СН2СН2СООН + 2 СО2 +2Н2 + Q
В качестве побочных продуктов может накапливаться бутиловый спирт, ацетон, этиловый спирт, уксусная кислота.
При масляно-кислом брожении сахар претерпевает те же превращения, что и при спиртовом и гомоферментативном молочнокислом брожениях, вплоть до образования пировиноградной кислоты. Пировиноградная кислота подвергается декарбоксилированию и при участии кофермента –А (КоА) расщепляется до ацетил-КоА, СО2, Н2. Две молекулы образовавщегося двууглеродного соединения конденсируются при участии фермента карболигазы. Из синтезированного четырехуглеродного соединения в сложном цикле последоваельных превращений образуется масляная кислота:
СН3СОСООН®СН3СНО + СО2; СН3СНО+ СН3СНО® СН3СН2СН2СООН
Маслянокислые бактерии представляют собой подвижные, крупные спорообразующие грамположительные палочки, строгие анаэробы.
В природе это брожение имеет положительное значение как звено в цепи многообразных превращений органических веществ. В пищевом производстве часто приносит значительный ущерб, так как может вызвать порчу картофеля и овощей, вспучивание сыров, порчу консервов (бомбаж), прогоркание молока, масла, увлажненной муки и т.д. Могут вызывать порчу квашеных овощей при замедленном молочнокислом брожении, образующаяся при этом масляная кислота придает продуктам острый прогорклый вкус и резкий неприятный запах.
Маслянокислое брожение применяют для производства масляной кислоты, которая представляет собой бесцветную маслянистую жидкость с неприятным резким запахом, а эфиры масляной кислоты отличаются прияным ароматом, как например: метиловый эфир имеет яблочный запах, этиловый- грушевый, амиловый – ананасовый. Их используют как ароматические вещества кондитерской, парфюмерной промышленности и при изготовлении фруктовых напитков.
Брожение пектиновых веществ. В тканях растений, овощей и фруктов пектины входят в состав клеточных стенок и срединных пластинок между отдельными клетками в виде труднорастворимого протопектина. Под действием пектолитических ферментов микроорганизмов, развивающихся в растительном пищевом сырье протопектин превращатся в растворимый пектин. Пектин разлагается с образованием галактуроновых кислот, углеводов (ксилозы, галактозы, арабинозы), метилового спирта и других веществ. Сахара сбраживаются бактериями по типу масляно-кислого брожения с образованием уксусной и масляной кислот, СО2 и Н2. Все эти процессы приводят к мацерации (распаду) пораженных объектов и другим видам повреждений. Разложение пектиновых веществ может быть выражено следующей схемой:
1. С46Н68О40 + nН2О® СНО (СНОН)4СООН + С6Н12О6 + С5Н10О5
Пектин галактуроновая к-та галактоза ксилоза
+ С5Н10О5 + СН3ОН +СН3СООН
арабиноза
2. С6Н12О6® СН3СН2СН2СООН + СН3СООН +СО2 +Н2+ Q
3. С5Н10О5® СН3СН2СН2СООН + СН3СООН +СО2 +Н2О+ Q
Продукты распада пектиновой кислоты (галактоза, арабиноза и др.) подвергаются сбраживанию или окислению разнообразными микроорганизмами. При анаэробных условиях они сбраживаются маслянокислыми бактериями относящимися к роду Клостридий. Пектиновое брожение наблюдается при мочке лубоволокнистых растений - льна, конопли, кенафа, джута, канатника и др. Для освобождения волокон клетчатки необходимо разложение пектина, что и происходит под действием пектиноразлагающих ферментов анаэробных бактерий.
Брожение клетчатки. Клетчатка (целлюлоза) является основным веществом клеточных стенок растений. Брожение клетчатки состоит из разложения ее в анаэробных условиях палочковидными споробразующими бактериями рода Клостридий. Они гидролизуют клетчатку до глюкозы в результате активности ферментов целлюлазы (до целлобиозы) и целлобиазы (до глюкозы) (см. уравнение 1-ниже). Глюкозу они сбраживают с образованием уксусной, масляной, молочной, муравьиной кислот, этилового спирта, СО2 и Н2 (уравнение 2 ниже):
1.(С6Н10О5)n + nН2О ®С12Н22О11 ® С6Н12О6;
2. С6Н12О6® СН3СООН +СН3СН2СН2СООН+ СН3СНОНСООН + С2Н5ОН +НСООН +СО2+ Q
Среди этих бактерий есть мезофилы и термофилы, которые накапливают различные органические кислоты.
Окислительные брожения. Окислительное брожение- это процесс окисления органических веществ (спиртов, углеводов) под действием ферментативной активности микроорганизмов не до конечных продуктов, как при дыхании, а до органических кислот и спиртов, как при брожении.
Уксуснокислое брожение. Это процесс окисления этилового спирта под действием ферментов уксусно - кислых бактерий до уксусной кислоты и воды. Окисление этилового спирта протекает в две стадии- сначала образуется уксусный альдегид, который далее окисляется до уксусной кислоты:
1. 2С2Н5ОН +О2® 2 СН3СНО + 2Н2О;
2. 2 СН3СНО+О2® 2 СН3СООН
Уксуснокислые бактерии бесспоровые палочки, грамотрицательные, строгие аэробы. Среди них есть подвижные и неподвижные формы. Уксусно-кислые бактерии относят к двум родам бактерий: Глюконобактер (Gluconobacter) и Ацетобактер (Acetobacter). Ацетобактер – перитрихи, способные окислять уксусную кислоту до СО2 и Н2О (переокисление).
Уксуснокислым бактериям свойственна изменчивость формы клеток, в неблагоприятных условиях могут образовывать толстые длинные нити, иногда раздутые уродливые клетки.
Уксуснокислое брожение лежит в основе получения уксуса для пищевых целей. Исходным сырьем может служить спиртовый раствор, разбавленное подкисленное плодово-ягодное или виноградное вино.
Лимонно-кислое брожение. Это процесс неполного окисления углеводов ферментативной активностью мицеллиальных грибов с образованием органических кислот (лимонной, щавелевой и др.). Окисление глюкозы в лимонную кислоту можно представить следующим суммарным уравнением:
2С6Н12О6+3О2® 2С6Н8О7 +4 Н2О
Химизм протекает по гликолитическому пути до образования пировиноградной кислоты. Пировинограданая кислота через ацетил-СoA включается в цикл Кребса (цикл ди- и трикарбоновых кислот), где происходит накопление яблочной, фумаровой, янтарной и лимонной кислот.
Наибольшее практическое значение имеет процесс получения лимонной кислоты с помощью гриба Аспергиллус Нигер (Aspergillus niger) в кислой среде. При изменении кислотности среды до нейтральной происходит накопление щавелевой кислоты.
Технические приемы биохимического получения лимонной кислоты в нашей стране были разработаны С.П. Костычевым В.С. Буткевичем. Используют поверхностный и глубинный (более перспективный) способы промышленного получения лимонной кислоты
Превращения белков - гниение. В метаболизме микроорганизмов азотсодержащие вещества подвергаются разнообразным превращениям. По случайно поверхностному сходству разные виды порчи пищевых продуктов нередко называют гниением. Однако гниение - это процесс глубокого разложения белковых веществ микроорганизмами.
Способность разлагать в той или иной степени белковые вещества свойственна многим микроорганизмам. Некоторые из них разлагают непосредственно белки, другие могут воздействовать только на более или менее простые продукты распада белковой молекулы, например на пептиды, аминокислоты и др.
Продукты разложения белков микробы используют для синтеза собственных органических веществ, а также в качестве энергетического материала.
Химизм разложения белковых веществ. Гниение - сложный, многоступенчатый биохимический процесс, характер и конечный результат которого зависят от состава разлагаемых белков, условий процесса и видов вызывающих его микроорганизмов.
Белковые вещества не могут непосредственно поступать в клетки микроорганизмов, поэтому использовать белки могут только те микроорганизмы, которые обладают протеолитическими ферментами — экзопротеазами, выделяемыми клетками в окружающую среду.
Процесс распада белков начинается с их гидролиза. Первичными продуктами гидролиза являются пептоны и пептиды. Они расщепляются до аминокислот, которые являются конечными продуктами гидролиза.
Образующиеся в процессе распада белков различные аминокислоты используются микроорганизмами или подвергаются ими дальнейшим изменениям, например дезаминированию, в результате чего образуются аммиак и разнообразные органические соединения. Процесс дезаминирования может происходить различными путями. Различают дезаминирование гидролитическое, окислительное и восстановительное.
Гидролитическое дезаминирование сопровождается образованием оксикислот и аммиака. Если при этом происходит и декарбоксилирование аминокислоты, то образуются спирт, аммиак и углекислый газ:
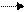 RCHNH
RCHNH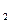 СOOН + НО RСНOНСООН + NН
СOOН + НО RСНOНСООН + NН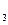 ;
;
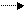 RСНNНСООН + НО RCHOH + NН+ CO;
RСНNНСООН + НО RCHOH + NН+ CO;
При окислительном дезаминировании образуются кетокислоты и аммиак:
RСНNНСООН + ½ O RСОCOOН+NНз
При восстановительном деэаминировании образуются карбоновые кислоты и аммиак:
 RСНNНСООН + 2Н RCHСООН + NН
RСНNНСООН + 2Н RCHСООН + NН
Из приведенных уравнений видно, что среди продуктов разложения аминокислот в зависимости от строения их радикала (R) обнаруживаются различные органические кислоты и спирты. Так, при разложении аминокислот жирного ряда могут накапливаться муравьиная, уксусная, пропионовая, масляная и другие кислоты, пропиловый, бутиловый, амиловый и другие спирты. При разложении аминокислот ароматического ряда промежуточными продуктами являются характерные продукты гниения: фенол, крезол, скатол, индол - вещества, обладающие очень неприятным запахом. При распаде аминокислот, содержащих серу, получается сероводород или его производные - меркаптаны (например, метилмеркаптан СНSН). Меркаптаны обладают запахом тухлых яиц, который ощущается даже при ничтожно малых концентрациях.
Образующиеся при гидролизе белка диаминокислоты могут подвергаться декарбоксилированию без отщепления аммиака, в результате чего получаются диамины и углекислый газ. Например, лизин превращается в кадаверин:
 NH
NH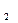 (CH)
(CH)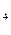 CHNHCOOOH декарбоксилаза NH(CH)
CHNHCOOOH декарбоксилаза NH(CH)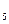 + CO
+ CO
Аналогично этому орнитин превращается в путресцин.
Кадаверин, путресцин и другие амины, образующиеся при гниении, часто объединяют под общим названием птомаины (трупные яды), некоторые из них обладают ядовитыми свойствами.
Дальнейшее превращение азотистых и безазотистых органических соединений, получающихся при распаде различных аминокислот, зависит от окружающих условий и состава микрофлоры. Аэробные микроорганизмы подвергают эти соединения окислению, так что они могут быть полностью минерализованы. В таком случае конечными продуктами гниения являются аммиак, углекислый газ, вода, сероводород, соли фосфорной кислоты. В анаэробных условиях не происходит полного окисления промежуточных продуктов распада аминокислот. В связи с этим кроме аммиака и углекислого газа накапливаются различные органические кислоты, спирты, амины и другие органические соединения, в числе которых могут быть вещества, обладающие ядовитыми свойствами, и вещества, придающие гниющему материалу отвратительный запах.
Возбудители гниения. Среди множества микроорганизмов, способных в той или иной мере разлагать белки, особое значение имеют микроорганизмы, которые вызывают глубокий распад белков - собственно гниение. Такие микроорганизмы принято называть гнилостными. Из них наибольшее значение имеют бактерии. Гнилостные бактерии могут быть спорообразующими и бесспоровыми, аэробными и анаэробными. Многие из них мезофилы, но есть холодоустойчивые и термостойкие. Большинство чувствительны к кислотности среды.
Наиболее распространенными и активными возбудителями гнилостных процессов являются Сенная и Картофельная палочки* - аэробные, подвижные, грамположительные, спорообразующие бактерии. Споры их отличаются высокой термоустойчивостью. Температурный оптимум развития этих бактерий 35 - 45°С, максимум роста - при температуре около 50 - 55°С; при температуре ниже 5°С они не размножаются.
* В соответствии с Международным кодексом номенклатуры бактерий сенная и картофельная палочки ввиду большого сходства объединены в один вид - Васillus subtilis.
Помимо разложения белков, такие бактерии способны разлагать пектиновые вещества, полисахариды растительных тканей, сбраживать углеводы.
Сенная и картофельная палочки широко распространены в природе и являются возбудителями порчи многих пищевых продуктов. Они вырабатывают антибиотические вещества, подавляющие рост многих болезнетворных и сапрофитных бактерий.
Бактерии рода Рsеudоmоnas - аэробные подвижные палочки, с полярным жгутиком, не образующие спор, грамотрицательные. Многие виды холодоустойчивы, минимальная температура их роста от - 2 до - 5°С, оптимум - около 20°С. Многие псевдомонасы помимо протеолитической обладают липолитической активностью; они способны сбраживать углеводы с образованием кислот, выделять слизь.
Развитие и биохимическая активность этих бактерий значительно тормозятся при рН ниже 5,5 и 5 - 6%-ной концентрации NаСl в среде. Псевдомонады широко распространены в природе, являются антагонистами ряда бактерий и плесеней, так как образуют антибиотические вещества. Некоторые виды Рsеudоmоnas являются возбудителями болезней (бактериозов) культурных растений, плодов и овощей.
Протей (Ргоtеus vulgаris) - мелкие грамотрицательные бесспоровыс палочки с резко выраженными гнилостными свойствами. Белковые субстраты при развитии в них протея приобретают сильный гнилостный запах. В зависимости от условий жизни эти бактерии способны заметно менять свою форму и размеры. Протей - факультативный анаэроб; сбраживает углеводы с образованием кислот и газов. Он хорошо развивается как при температуре 25 °С, так и при 37°С, прекращая размножаться лишь при температуре около 5°С, однако может сохраняться и в замороженных продуктах.
Характерной особенностью протея является его очень высокая подвижность. Это свойство лежит в основе метода выявления протея на пищевых продуктах и отделения его от сопутствующих бактерий. Некоторые виды протея выделяют токсические для человека вещества.
Клостридиум путрификум (Сlоstridium putrificum) - анаэробная подвижная, спорообразующая палочка. Относительно крупные споры ее располагаются ближе к концу клетки, которая при этом приобретает сходство с барабанной палочкой. Споры довольно термоустойчивы. Углеводы эта бактерия не сбраживает. Белки разлагают с образованием большого количества газов (NН, Н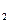 S). Оптимальная температура развития 37 – 43 °С, минимальная 5 °С.
S). Оптимальная температура развития 37 – 43 °С, минимальная 5 °С.
Клостридиум спорогенес (Сlоstridium sporogenes) - анаэробная подвижная спороносная палочка. Споры термоустойчивы, в клетке они расположены ближе к ее концу. Характерным является очень быстрое (в течение первых суток роста) образование спор. Эта бактерия сбраживает углеводы с образованием кислот и газа, обладает липолитической способностью. При разложении белков обильно выделяется сероводород. Оптимальная температура развития 35-40 °С, минимальная-около 5°С.
Оба вида клостридий известны как возбудители порчи баночных консервов (мясных, рыбных и др.).
Практическое значение процессов гниения. Гнилостные микроорганизмы наносят нередко большой ущерб народному хозяйству, вызывая порчу ценнейших и богатых белками продуктов питания, например мяса и мясопродуктов, рыбы и рыбопродуктов, яиц, молока и др. Но эти микроорганизмы играют большую положительную роль в круговороте веществ в природе, минерализуя белковые вещества, попадающие в почву, воду.
Нитрификация. Процесс последовательного окисления аммиака до азотистой и азотной кислот называется нитрификацией, а возбудители его—нитрифицирующими бактериями. Сущность этого процесса была раскрыта и изучена С. Н. Виноградским.
Работами С. Н. Виноградского установлено, что процесс нитрификации происходит в две фазы, каждая из которых обусловлена деятельностью специализированных аэробных бактерий. Возбудители первой фазы — нитрозные бактерии-окисляют аммиак до солей азотистой кислоты (нитритов). Возбудители второй фазы - нитратные бактерии - окисляют соли азотистой кислоты в соли азотной кислоты (нитраты).
Нитрифицирующие бактерии относятся к хемоавтотрофам. Они живут в почве, природных водоемах, обогащая их нитратами - лучшим источником азотистого питания для растений.
Денитрификация. Процесс восстановления нитратов до молекулярного азота называется денитрификацией, а бактерии, осуществляющие его,— денитрифицирующими бактериями.
Денитрифицирующие бактерии - факультативные анаэробы. В аэробных условиях в процессе их дыхания конечным акцептором водорода является кислород. В анаэробных условиях в качестве акцептора водорода используются нитраты (некоторые используют нитриты), которые и восстанавливаются до молекулярного азота. Многие микроорганизмы восстанавливают нитраты только до нитритов.
Денитрифицирующие бактерии широко распространены в природе. Они живут в почве, природных водах. Деятельность денитрифицирующих бактерий в почве отрицательна, особенно при анаэробных условиях, так как азот нитратов, усваиваемый растениями, переходит в неиспользуемый ими свободный азот.
Фиксация молекулярного азота. Некоторые бактерии способны фиксировать свободный атмосферный азот, т. е. переводить его в связанное состояние. Они восстанавливают азот до аммиака; часть его используется самими микроорганизмами, а часть выделяется в окружающую среду.
Одни азотфиксирующие (азотусваивающие) бактерии живут свободно в почве и в воде; другие — в симбиотическом сожительстве с растениями, преимущественно бобовыми, Бактерии поселяются в бородавчатых вздутиях — клубеньках корней этих растений. Отсюда произошло и название этих бактерий — клубеньковые. Необходимую для фиксации азота энергию они получают в процессе окисления безазотистых органических соединений.
Среди свободно живущих азотфиксирующих бактерий наибольшее значение имеют следующие: аэробные бактерии рода азотобактер (Аzоtоbасtег) — слегка приплюснутые кокки, часто объединенные попарно, имеющие слизистую капсулу; анаэробные бактерии, открытые С. Н. Виноградскнм, Clostridium pasterianum—подвижные спорообразующие палочки, способные сбраживать углеводы по типу маслянокислого брожения, которое и служит этим бактериям источником энергии для связывания молекулярного азота.
Азотфиксирующие бактерии имеют большое значение. В результате их деятельности почва обогащается доступной для растений формой азотистого питания.
В практике сельского хозяйства в качестве бактериального удобрения используют препараты азотфиксирующих бактерий—азотобактерин (из культур азотобактера) и нитрагин (из культур клубеньковых бактерий).
Окружающая среда, кожные покровы и слизистые оболочки верхних дыхательных путей и других органов человека и животных заселены многочисленными группами микроорганизмов. Их жизнь тесно связана с жизнью хозяина (макроорганизма). В большинстве случаев сложившийся симбиоз является выгодным для микро- и макроорганизмов. Например, пищеварительный тракт человека и животных заселен микрофлорой которая питается за счет хозяина и одновременно помогает организму лучше переваривать пищу и корма, создавая при этом неблагоприятные условия для жизнедеятельности других микробов (мутуализм).
В отдельных случаях микроорганизм обладает способностью проникать во внутреннюю среду организма-хозяина и размножаться, принося ему вред (паразитизм). Поскольку микроорганизмы-паразиты приспособились в процессе эволюции к обитанию в определенных тканях организма животных и людей, их присутствие и размножение приводит к повреждению клеток, нарушению функции органов и систем организма, а следовательно, к заболеваниям.
|
|
|
|
|
Дата добавления: 2013-12-13; Просмотров: 1278; Нарушение авторских прав?; Мы поможем в написании вашей работы!