КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Хромосомные и геномные мутации
Различные клетки одного организма и различные особи одного вида обладают, как правило, одинаковым числом хромосом в наборе, за исключением гаметных клеток, которые содержат вдвое меньше хромосом по сравнению с соматическими клетками.
Число гомологичных хромосом и порядок генов в них также, как правило, совпадают в различных клетках и у разных представителей одного вида. Из этих правил существуют исключения, известные как хромосомные мутации (перестройки или аберрации).
Хромосомные мутации (хромосомные аберрации) - следующие после одного или нескольких хромосомных разрывов внутрихромосомные или межхромосомные перемещения или потери хромосомных сегментов, имеющие генетические последствия.
Хромосомные аберрации вызваны перемещением генетического материала, приводящим к изменению структуры хромосом в пределах кариотипа. В такие перестройки могут быть вовлечены участки одной хромосомы или разных – негомологичных - хромосом.
В соответствии с этим критерием выделяют аберрации внутрихромосомные и межхромосомные.
К внутрихромосомным относятся: нехватки, дупликации, инверсии.
1) Нехватки могут быть концевыми (их называют дефименси), когда происходит потеря одного или двух концевых участков хромосомы (рис.а, б,в) и срединными – делеции, возникающие вследствие потери небольшого внутреннего хромосомного или хроматидного сегмента в результате двух разрывов (рис.).
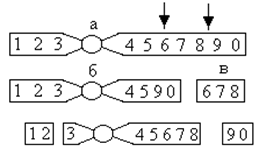 |
Рисунок. Дефименси (а – исходная хромосома, б - потеря одного и в – двух терминальных участков).
Рисунок. Делеция (а – исходная хромосома, б – хромосома с делецией, в – ацентрический фрагмент).
В случае дефименси после потери двух терминальных участков открытые концы хромосомы могут соединяться, образуя кольцеобразную хромосому.
Фрагмент хромосомы элиминируется в митозах.
В случае делеции плечо хромосомы с большой нехваткой становится короче, а выпавший сегмент может образовывать ацентрическое кольцо, элиминирующееся при делении клетки. Так как вследствие делеций теряются участки хромосом, у гетерозигот по этим перестройкам наблюдаются характерные нарушения конъюгации гомологов. Более длинная нормальная хромосома образует петлю на участке, соответствующем делеции.
Делеция доминантного аллеля в гетерозиготе приводит к проявлению рецессивного аллеля. Это явление называется псевдодоминированием.
2) Дупликация – разновидность хромосомных перестроек, когда некоторый участок хромосомы в гаплоидном в наборе представлен два раза. Гетерозиготные по дупликации особи несут две дозы дуплицированных генов, гомозиготные – четыре. Дупликация происходит вследствие как минимум трех разрывов и последующей нехватки (делеции) в одной хромосоме и транслокации выпавшего участка в другую хромосому (рис.)
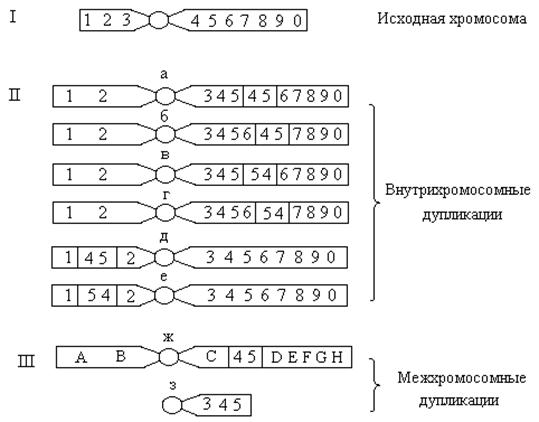 |
Рисунок. Типы дупликаций.
Если сегмент перемещен в гомологичную хромосому, дупликация называется внутрихромосомной. Внутрихромосомная дупликация может быть смежной (тандемной, рис., а) или несмежной (рис., б), внутриплечевой (рис., г), межплечевой (рис., д, е). Если в дуплицированном сегменте сохраняется порядок генов исходной хромосомы, дупликация называется прямой (рис., а, в, д), при переворачивании дуплицированного сегмента на 1800 – обратной или дупликацией с инверсией (в, г, е). Если дуплицированный сегмент локализован в негомологичной хромосоме (ж) или присутствует в хромосомном наборе в форме центрического фрагмента (з), дупликация называется межхромосомной.
Известны случаи многократных повторений (мультипликаций) какого-либо участка, – амплификации.
Главной причиной множественных повторов участков генетического материала является неравный кроссинговер, при котором разрывы происходят не в строго идентичных, а в разных местах несестринских гомологичных хроматид, вследствие чего и обмен между этими хроматидаим происходит неравными сегментами: в хроматиде, отдавшей меньшей сегмент, образуется дупликация; в хроматиде, вчленившей этот меньший сегмент и отдавшей большой – делеция.
Дупликации играют существенную роль в эволюции генома, так как они создают дополнительные участки генетического материала, функция которых может быть изменена в результате мутаций и последующего естественного отбора.
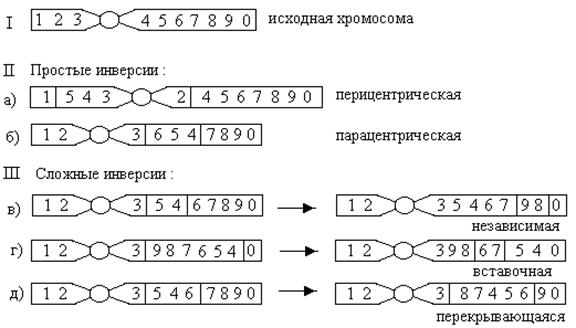 |
3) Инверсия – внутрихромосомная мутация, возникающая в результате двух или большего числа разрывов и перевертывания участка хромосомы на 1800 (рис.). Инверсии часто встречаются как хромосомные перестройки, но могут быть и хроматидные инверсии. Если инвертированный сегмент включает центромерную область, инверсия называется перицентрической (симметричной), если нет – парацентрической (асимметричной).
Рисунок. Типы инверсий.
Инверсия называется простой, если инвертирован один сегмент хромосомы, сложной, если инвертированы два сегмента. Сложная инверсия может быть независимой, если второй инвертированный сегмент лежит вне первого, вставочной, если вторая инверсия происходит внутри первоначального инвертированного сегмента, и перекрывающейся, если второй инвертируемый сегмент одним концом захватывает первую инверсию.
Инверсия в гомозиготном состоянии часто связана с летальным эффектом, поэтому ее наблюдают большей частью в гетерозиготах. Другое важное следствие инверсии – подавление кроссинговера. При конъюгации гомологичных хромосом в мейозе гетерозигот по инверсии наблюдаются петли. Если в такой петле (в инвертируемом участке) произойдет одинарный кроссинговер, то в случае парацентрической инверсии возникает одна, хроматида с двумя центромерами, (дицентрическая хромосома), которые ее порвут при расхождении. Вторая хроматида будет без центромеры (аценрическая хромосома), она элиминируется в процессе мейоза. Таким образом, из четырех гамет жизнеспособными окажутся только две. Вследствие этого инверсия рассматривается как косвенный подавитель кроссинговера.
Таким образом, гетерозиготные инверсии служат факторами изоляции и способствуют эволюционной дивергенции новых форм, образующихся в пределах вида.
В то же время инверсии – широко распространенный путь эволюционного преобразования генетического материала. Например, кариотипы человека и шимпанзе при почти подобном геноме имеют различное число хромосом: Homo sapiens – 2n = 46, Anthropopithecus pan – 2n = 48. Хромосома 2 человека содержит большую часть материала, гомологичного дополнительной паре хромосом шимпанзе. Кроме того, различая касаются четырех хромосом (аутосом): 4-ой, 5-ой, 12- ой, 17-ой, в которых произошли перицентрические инверсии.
К межхромосомным аберрациям относятся траспозиции и транслокации. Под термином «транслокация» в широком смысле понимается любое перемещение хромосомных сегментов в хромосомном наборе. Если перемещается один сегмент, транслокация называется простой, или односторонней, если меняются местами два сегмента – взаимной, или реципрокной. Транслокация может происходить внутри одной и той же хромосомы (внутрихромосомная транслокация), в пределах одного или двух ее плеч. Межхромосомной транслокацией называется перемещение сегментов между двумя гомологичными (гомосомальная или сестринская) или негомолочными (гетеросомальная или несестринская транслокация) хромосомами. Если порядок генов по отношению к центромере в траслоцированном сегменте остается исходным, транслокация называется эуцентрической, если сегмент инвертирует – дисцентрической.
В узком смысле под траслокацией понимается обмен сегментами между двумя негомологичными хромосомами – взаимные или реципрокные транслокации. Это наиболее распространенный тип транслокаций. В этом случае может образоваться две новые моноцентрические хромосомы или дицентрическая хромосома с одним или двумя ацентрическими фрагментами. Реципрокные транслокации проявляются, как правило, в гетерозиготном состоянии. В потомстве этих гетерозигот появляются гомозиготы по транслокации, не связанные с летальным эффектом.
Крупные транслокации изменяют морфологию хромосом и картину их конъюгации в мейозе (кресты, кольца, восьмерки из хромосом), межхромосомные – изменяют группы сцепления, что обнаруживается при гибридологическом анализе. Характер конъюгации траслоцированных хромосом меняется, образуется фигура креста. Вблизи точек разрывов конъюгация затрудняется и в этих участках подавляется кроссиговер.
Таким образом, у гомозигот по транслокациям изменяется характер сцепления генов. В гетерозиготе по траслокациям гены, принадлежащие к негомологичным хромосомам, наследуется как принадлежащие к одной группе сцепления, так и полностью функциональными оказываются те гаметы, которые несут родительские сочетания хромосом. Участок хромосомы может также изменять свое положение и без реципрокного обмена, оставаясь в той же хромосоме или включаясь в какую-то другую. Транслокации такого типа иногда называю транспозициями.
Транспозиция – перемещение фрагмента генетического материала в пределах одной хромосомы (инсерция) или между разными хромосомами (вставка), происходящее вследствие трех разрывов. Происходит при участии мобильных диспергированных генетических элементов (МДГ – элементов) – нуклеотидных последовательностей, способных перемещаться с одного места на другое – транспазонов. В 1947 – 1950 гг. американка Б. Мак Клинток, изучая генетику окраски зерна кукурузы, выявила мутации и хромосомные перестройки под действием определенных генетических элементов, перемещающихся в пределах генома. Транспазонные элементы были открыты при нахождении вставок нового материала в границах бактериальных оперона. Такие вставки локализуются в середине гена и препятствуют его транскрипции. Последовательности нуклеотидов, способные вклиниваться в разных участках генома клинической палочки (E. сoli), называются IS-элементами (от англ. Insertion sequences – вставные последовательности). Их генетические функции связаны лишь со способностью к транспозиции. Большинство IS-элементов имеют ген, контролирующий синтез фермента транспозазы, ответственного за их перемещение. Таким образом, вместе с плазмидами транспазоны прокариот определяют подвижность генов хозяина и компенсируют отсутствие полового процесса. У эукариот траспазоны обнаружены у дрожжей и дрозофил. Их особенность – наличие концевых повторов. Вне генома транспазоны про- и эукариот функционировать не могут. Следовательно, основными функциями МДГ-элементов являются:
1. обеспечение рекомбинации генетического материала;
2. создание условий непостоянства генома;
3. вызывают генные мутации и хромосомные перестройки;
4. обеспечение переноса нуклеотидных последовательностей, находящихся рядом, в новое место на хромосоме.
Перенос генов МДГ- один из факторов эволюции. Кроме обычных путей передачи наследственной информации от родителей потомкам, существует еще и горизонтальный перенос генетического материала между отдельными видами, поэтому разнообразие генетической изменчивости может увеличиваться.
Таким образом, изменение в структуре хромосом (хромосомные мутации) могут затрагивать число генов в хромосомах (инверсии и транслокации).
К числу хромосомных некоторые авторы (Ф. Айала, 1984 и др.)относят и геномные мутации, связанные с изменением количества хромосом в геноме. При изменениях такого рода в одних случаях (смещения и разделения) общее количество наследственного материала остается неизменным, а в других (анеуплоидия, моноплоидия, полиплоидия) - изменяется.
1. Центрическое слияние. Две негомологичные хромосомы сливаются в одну. Это неизбежно приводит к утрате центромеры.
2. Центрическое разделение. Одна хромосома разрывается на две. При этом должна образоваться новая центромера; в противном случае хромосома без центромеры утрачивается при клеточном делении.
3. Анеуплоидия – гетероплоидия, т.е. уменьшение или увеличение числа хромосом, некратное основному числу хромосом вида. Термины нуллисомик и моносомик относятся к организмам, содержащим соответственно на одну пару хромосом и на одну хромосому меньше нормы. Термины трисомик, тетрасомик и т.д. означают, что в хромосомном наборе присутствуют соответственно одна, две и т.д. лишние хромосомы.
4. Моноплоидия и полиплоидия. При этом число наборов негомологичных хромосом отличается от двух. Большинство эукариотических организмов диплоидны, т.е. несут по два набора негомологичных хромосом в каждой соматической клетке и по одному набору – в гаметах. Наряду с этим есть организмы, которые в норме моноплоидны, т.е. содержат по одному набору хромосом. Например, у пчел самцы моноплоидны и развиваются из неоплодотворенных яиц партеногенетически, а самки диплоидны и развиваются из оплодотворенных яиц.
Полиплоидные организмы имеют более двух наборов негомологичных хромосом. Организм называется триплоидным, если он имеет три набора хромосом, тетраплоидным, если он несет четыре набора и т.д. Наиболее распространены полиплоидные организмы, у которых число хромосомных наборов в клетке кратно двум: тетраплоиды, гексаплоиды и октоплоиды, содержащие соответственно четыре, шесть и восемь хромосомных наборов. Полиплоидия очень распространена в некоторых группах растений, но редко встречается у животных.
Индуцирующие факторы, описанные выше, могут вызывать перечисленные повреждения на хромосомном уровне. Это приводит к хромосомным мутациям: транслокациям, инверсиям, делециям, дефименсам, дупликациям. Хромосомные мутации регистрируются под световым микроскопом. Мутации этого типа сильнее влияют на отклонения от нормального фенотипа, а потери больших участков хромосом приводят к летальным исходам еще во внутриутробном периоде. Поэтому такие мутации выявляются чаще при анализе кариотипа у лиц с врожденными пороками развития или у спонтанно абортированных эмбрионов.
Частота хромосомных и геномных мутаций у человека и методы ее оценки.
Частота хромосомных и геномных мутаций в популяциях человека намного выше, чем генных. Их наличие в половых клетках объясняет высокую летальность гамет и зигот на ранних стадиях развития (до 50-70%). Таким образом, у человека действует довольно эффективныйпренатальныйи постнатальный отбор. Спонтанные аборты составляют около 15% всех беременностей, мертворождения – 2%, а детская смертность – 2-3 %. Жесткому давлению отбора подвергаются, прежде всего, хромосомные и геномные мутации, о чем свидетельствует прогрессирующее уменьшение их частоты в ряду: эмбрион – плод – новорожденный. Однако 3-4 ребенка из каждой 1000 новорожденных страдают хромосомными болезнями (см. гл.). Большинство таких болезней являются спорадическими, то есть возникают вновь вследствие геномных (или хромосомных) мутаций в гаметах здоровых родителей или в зиготах при первых их митотических делениях.
По прогнозам ученых, в будущем развитие общества и медицины приведет к тому, что жизнеспособность и плодовитость мутантных организмов будет приближаться к таковым у здоровых людей. В этом случае, в соответствии с принципами популяционной генетики, количество больных будет возрастать из поколения в поколение. В этой связи изучение динамики частоты хромосомных и геномных мутаций у человека является достаточно актуальным.
Установлено, что для заболеваний, протекающих тяжело и формирующихся внутриутробно (например, хромосомных болезней), новые мутации, возникающие в половых клетках здоровых родителей, составляют от 79 до 94 %, а семейные формы, т.е. унаследованные от больных родителей - только 6-21 %.
При изучении частоты возникновения хромосомных дефектов в популяциях человека в последнее время установлено, что:
1. В человеческих популяциях достаточно часто (чаще, чем у млекопитающих животных) встречаются структурные перестройки (инверсии, транслокации).
2. При изучении частоты первичных мутационных событий (нерасхождения, перестройки хромосом) необходимо проводить кариологическое обследование супружеских пар, особенно при изучении спонтанных абортов. При кариотипировании материала спонтанных абортов многие случаи аномалий будут относиться к унаследованным, а не первично возникшим событиям.
3. Существенным признаком популяции является возраст родителей, особенно матери, т. к. твердо установлена зависимость частоты анеуплоидий по многим хромосомам от возраста матерей (табл.).
4. Поскольку частота хромосомных аномалий неодинакова на разных стадиях онтогенеза (спонтанные аборты, пренатальная смертность, живорождения, разные возрастные группы),то учет хромосомных аномалий в популяции должен быть на всех стадиях с одновременной оценкой частот разных исходов беременности. Различная частота спонтанных абортов, мертворождений и живорождений в популяциях может быть обусловлена другими (социально-экономическими, биологическими) факторами, не связанными с хромосомными и геномными мутациями.
5. Среди всех больных с хромосомными заболеваниями, не менее25% являются больными с мозаицизмом. У многих из них нетяжелые формы заболевания; у них рождаются дети, что в свою очередь повышает процент хромосомных аномалий в популяциях. Указанный процент мозаицизма свидетельствует о высокой частоте возникновения мутаций на ранних этапах развития эмбриона, т. е. в соматических клетках. В связи с этим при популяционных обследованиях необходимо оценивать мозаицизм у супружеских пар и пробандов, чтобы определить частоту первичных событий в зародышевых клетках.
Изучение распространения хромосомной патологии как части общего груза наследственных болезней в разных группах населения позволяет оценить роль хромосомной патологии в общей патологии человека, а также частоту возникновения хромосомных и геномных мутаций. При оценке параметров распространения хромосомных аномалий в популяциях человека используют данные цитогенетических исследований, проведенных на материале спонтанных абортов, мертворожденных и у новорожденных детей.
Результаты по первым двум группам характеризуют хромосомные аномалии как этиологический фактор внутриутробной летальности. Реальная картина хромосомных аномалий в популяциях основывается на результатах цитогенетического исследования у новорожденных детей. Для оценки частоты возникновения мутаций этого недостаточно. Необходимо также иметь сведение о внутриутробных летальных эффектах хромосомных и геномных мутаций.
|
|
Дата добавления: 2014-01-04; Просмотров: 4685; Нарушение авторских прав?; Мы поможем в написании вашей работы!