КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
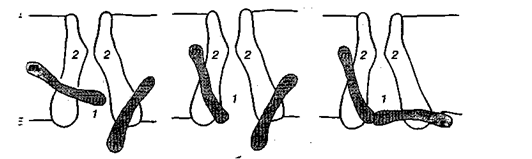
Мал. 2. Механізм виникнення двофазного потенціалу дії: а, б — відвідні електроди
|
|
|
|
Залежно від умов відведення розрізняють двофазний та однофазний ПД. Перший виникає, коли збудження, що поширюється нервовою чи м'язовою клітиною, проходить послідовно спочатку під одним, а потім під другим відвідним електродом. Щоб отримати однофазний ПД, треба ділянку тканини під другим, що знаходиться далі від подразнювальних, електродом зруйнувати. Тоді збудження не досягає другого електрода і на екрані осцилографа ми бачимо лише першу фазу ПД.
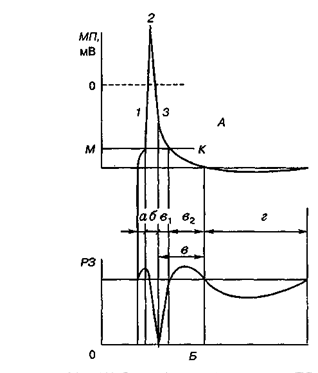
При внутрішньоклітинному відведені можна кількісно охарактеризувати зміни МПС при генерації ПД. У типовому внутрішньоклітинному ПД розрізняють пік (верхівку), висхідну, низхідну фази і слідові потенціали. Встановлено, що під час висхідної фази ПД відбувається не просто зникнення трансмембранної різниці потенціалів (деполяризація мембрани), а виникає різниця потенціалів протилежного знака (овершут), внутрішня частина мембрани в ділянці генерації ПД стає на певний час зарядженою позитивно відносно зовнішньої частини мембрани, зарядженої негативно.
Під час низхідної фази ПД відновлюється початкове значення МП (реполяризація мембрани). Тривалість піку ПД різних збудливих тканин варіює від 0,5 до 3 мс. Зміни поляризації мембрани, які спостерігаються після завершення високовольтної частини ПД (піку), називають слідовими потенціалами. Розрізняють два види слідових потенціалів — слідову деполяризацію (слідовий негативний потенціал) і слідову гіперполяризацію (слідовий позитивний потенціал), амплітуда яких не перевищує кількох мілівольт (5-10% рівня піку), а тривалість буває різною — від кількох мілісекунд до десятків секунд.
Іонний механізм генерації ПД. В основі генерації ПД лежать зміни проникності клітинної мембрани, що виникають послідовно за часом. У момент подразнення провідність мембрани для Nа+ зростає. Nа+ починає надходити в клітину, знижуючи внутрішній негативний заряд мембрани, тобто виникає деполяризація клітинної мембрани. При досягненні мембраною певного — критичного — рівня деполяризації виникає лавиноподібний потік Nа+ всередину клітини, що зумовлює появу висхідного коліна ПД. У цей момент gNа зростає у 100 разів порівняно з: станом спокою і стає у 20 разів більшої за gк. І якщо gNа збільшується далі, то заряд мембрани в місці подразнення змінюється — виникає реверсія знака МП. Проте під час збудження позитивне значення ПД (овершут ) не може бути білішим за натрієвий рівноважний потенціал, (ЕNа), який становить +65мВ. Звичайно овершут не перевищує + 30 мВ.
|
|
|
Існує принаймні дві причини цього явища: 1) підвищення gNа не триває довго, коли досягнуто піка ПД (протягом 1 мс), gNа починає різко падати, так що мембрана не встигає перезарядитися до + 65 мВ; 2) в той самий час, приблизно в межах 1 мс після початку збудження, відбувається повільніше зростання gк, що зумовлює фазу реполяризації мембрани.
Натрієва провідність досягає максимуму менш ніж за 1 мс після початку ПД, а потім різко знижується, зникаючи зовсім протягом 0,5 мс, хоча деполяризація мембрани ще зберігається. Це явище називають натрієвою інактивацією, яка існує доти, доки триває деполяризація мембрани. Натрієва активаційна система відновлюється, коли МПС повертається до початкового рівня або навіть при певній гіперполяризації мембрани. На відміну від gNа, gк не інактивується і навіть зростає доти, доки існує деполяризація мембрани.
Зміни збудливості клітини під час збудження. Під час натрієвої інактивації будь-яка деполяризація мембрани абсолютно неефективна, тобто клітина в цей час незбудлива. Це абсолютний рефрактерний період, який триває 1—2 мс. Потім протягом кількох мілісекунд після закінчення абсолютної рефрактерності нове збудження можна викликати лише більшою силою подразнення, ніж перший ПД. Значення ПД в цей час також зменшене, оскільки натрієва система не повністю відновлена після інактивації під час першого ПД. Цей відрізок часу, протягом якого відбувається відновлення збудливості клітини, називають відносним рефрактерним періодом.
|
|
|
Абсолютний рефрактерний період обмежує максимальну частоту генерації ПД живою клітиною. Оскільки у більшості нервових клітин П Д триває близько 2 мс, то максимально можлива частота генерації ПД у них буде 500 за 1 с. Проте є клітини з ще коротшим рефрактерним періодом, які мають частоту генерації до 1000 за 1 с. Зазвичай частота генерації ПД в ЦНС не перевищує кількох десятків ПД за 1 с.
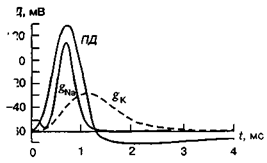
Мембранна провідність під час розвитку ПД у гігантському аксоні кальмара
Під час слідової деполяризації збудливість клітини підвищується (фаза екзальтації), а під час слідової гіперполяризації — знижується, оскільки значення МПС у цей час зростає, при цьому збільшується і критичний (пороговий) рівень деполяризації мембрани.
У живій клітині існують дві системи руху іонів крізь мембрану. Одна з них здійснюється за градієнтом концентрації іонів і не потребує витрати енергії — це пасивний іонний транспорт. Він відповідає за підтримання МПС і виникнення ПД, здійснюється через іонні канали. Друга система руху іонів крізь мембрану відбувається проти концентраційного градієнта і полягає у "викачуванні" Na+ з цитоплазми і "закачуванні" К+ всередину клітини за допомогою і онних насосів. Цей механізм можливий тільки з витратами енергії і називається активним іонним транспортом.
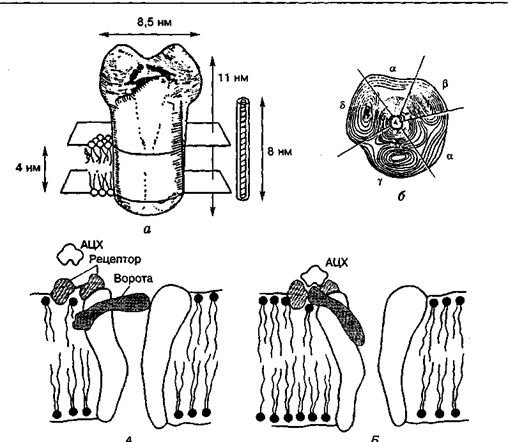
Мал. 3. Холінорецептор електричного органа риби:
тривимірне зображення рецептора збоку (а) і згори (б); е — активація рецептора ацетилхоліном (АЦХ); А — ворота зачинені;Б—ворота відчинені
Функції іонних каналів. У клітинній мембрані існують пори, або канали. Терміном "канал" позначають той шлях, яким іони проходять крізь мембрану за електрохімічним градієнтом. У клітинній мембрані існує кілька різних іонних каналів: селективних та неселективних. Перші пропускають лише якийсь певний іон: Na+, К+ або Са2+, а другі — кілька іонів. Крім того, селективні канали поділяють на потенціалозалежні, потенціалонезалежні (хемочутливі) і канали витоку.
|
|
|
Іонні канали
| Селективні (дляNa+, К+ або Са2+) | Неселективні | ||
| Потенціалонезалежні | Потенціалозалежні (хемочутливі) | Канали витоку | |
Потенціалозалежні канали. Провідність цих каналів контролюється рівнем МПС, тобто ці канали відкриваються (активуються) і закриваються (інактивуються) у відповідь на зміни МПС. Потенціалозалежні канали функціонують в електрозбудливих мембранах і беруть участь у генерації ПД. Електрична збудливість пов’язана з існуванням у таких каналах особливого молекулярного пристрою — воріт, відкривання і закривання яких визначається електричним полем мембрани.
Деполяризація мембрани, мВ
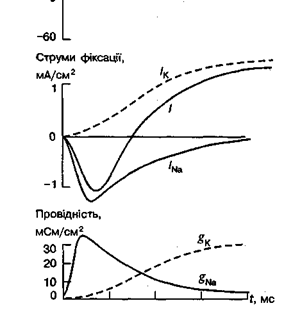
Мал. Іонні струми і зміна провідності при зміні МП у гігантському аксоні кальмара: зверху показано деполяризаційне зрушення потенціалу від -60 до 0 мВ. Верхні криві показують підсумковий струм фіксації I та його компоненти Iк і INa; нижні криві показують часовий перебіг змін мембранної провідності gNа і gк, розрахованих за рівнем струмів фіксації
Ворота можуть перебувати тільки у двох положеннях: повністю відчиненими чи повністю зачиненими, тому провідність поодинокого іонного каналу є сталою. Через відкриті канали іони рухаються за концентраційним і електрохімічним градієнтами. Ці потоки іонів зумовлюють зміну МПС, що, в свою чергу, змінює середню кількість відкритих каналів і відповідно рівень іонних струмів.
Для дослідження функції іонних каналів застосовують різні методи. Один із них це метод фіксації напруги на мембрані за допомогою якого на мембрані штучно підтримується будь-який потрібний потенціал. Функцію каналів відбивають іонні струми, які в цей час проходять крізь мембрану.
Потенціалозалежними є натрієві, калієві і кальцієві канали. Натрієві і кальцієві канали відповідають за ранній (початковий) струм вхідного (в клітину) напрямку, а калієві — за пізній (відставлений) струм вихідного (з клітини) напрямку.
|
|
|
Рівень gNа і gк відбиває кількість одночасно відкритих натрієвих і калієвих каналів. Як можна побачити, gNа за частки мілісекунди досягає максимуму, а потім повільно знижується до початкового рівня. Це пояснюється наявністю у натрієвих каналах двох типів воріт — швидких активаційних і повільних інактиваційних. Отже, початкове збільшення gNа пов'язане з відкриванням перших воріт (процес активації), а наступне повільне зниження gNа, що відбувається під час деполяризації мембрани, зумовлене: закриванням других воріт (процес інактктивації).
Натрієвим каналам властива досить висока, але не абсолютна вибірковість Nа+, оскільки більшою або меншою вони є проникними також для іонів літію, талію і навіть деяких органічних катіонів (гідразин, амоній). Отвір натрієвого каналу має прямокутну форму розмірами 0,3х0,3 нм. Вузька частина каналу, що є селективним фільтром, дуже коротка і має одиничний негативний заряд. Цей заряд притягує катіони, відштовхує аніони, забезпечуючи вибіркову проникність для Nа+ і подібних до нього катіонів.
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 845; Нарушение авторских прав?; Мы поможем в написании вашей работы!