КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Лекция 2. Логическое продолжение: от потенциала воспроизводства – к схеме воспроизводства
|
|
|
|
Логическое продолжение: от потенциала воспроизводства – к схеме воспроизводства
Потенциал воспроизводства велик, поэтому составляющих много –велика надстройка (которая собственно и является живой системой) за счет потенциала воспроизводства
+ в сравнении с равновесием (без воспроизводства), где в обе сторону скорости одинаковы и потребление энергии извне не происходит, это должен быть «маховик» воспроизводства как движение в определенном направлении (в сторону синтеза воспроизводимых составляющих, распад происходит самопроизвольно или некоторым образом предотвращается/компенсируется)
В результате воспроизводство существенно невоспроизводимого поведения (как при жизнедеятельности) должно включать:
собственно синтез составляющих как их химическое воспроизводство
+ воспроизводство способности к непредсказуемому поведению (вплоть до свободы воли), которое требует специального механизма (автоматически не происходит или очень мал о)
+ энергетическое обеспечение первого и второго (если его нет, то первый и второй остановятся, как направленные процессы)
(!) составляющих много (именно в силу того, что потенциал воспроизводства велик) и по смыслу эволюционного процесса все важны:
последовательное добавление составляющих способно конкурировать с другими системами (в которых составляющие не добавляют), если каждая добавка существенно увеличивает эффективность жизнедеятельности (воспроизводство с добавкой позволяет опередить воспроизводство без добавки для исходной схемы и у отличающихся конкурентов)
при большом числе составляющих и сложной организации их взаимодействий (когда каждая существенна или принципиально важно) особую значимость приобретает представление того, как они между собой взаимодействуют:
(О) схема воспроизводства – описание взаимодействия составляющих живой системы, результатом которого является воспроизводство системы как единого целого (т.е. совокупности всех составляющих) = как составляющие превращаются так, чтобы получить все из всех (с использованием внешних ресурсов) + хорошо иметь степени свободы «на вырост»
Следствия:
1. для получения (производства) каждой составляющей должно быть взаимодействие (процесс), представляемое в схеме воспроизводства
2. получение каждой составляющей должны быть связано с другими (схема не должна «разваливаться» и от нее не должны «отваливаться куски», иначе будет разделение схем воспроизводства или некоторые составляющие не будут нужны для воспроизводства схемы как совокупности других)
3. если есть процессы получения (производства) одной или малого числа составляющих, которое осуществляют многие составляющие (и они специфически необходимы именно для этого получения, участвуя только в нем) то, значит, должны быть и процессы получения многих составляющих, которое обеспечивает относительно малое число составляющих
… (попробуйте дополнить)
(П) схема воспроизводства на молекулярном уровне (универсальный биологический механизм):
(О) репликация:
– синтез ДНК из нуклеотидов на ДНК (осуществляющей долговременное хранение всей информации) = одна составляющая получается из одной (та же самая, если не учитывать возможные ошибки) с участием нескольких участников полимеразного процесса, которых относительно мало в сравнении с общим числом составляющих в схеме воспроизводства
(О) транскрипция:
– синтез мРНК (для кратковременного хранения информации) из нуклеотидов на ДНК = многие составляющие получаются из одной с участием относительно малого числа участников полимеразного процесса
+ есть синтез других РНК
(О) трансляция:
– синтез белков (перевод информации в исполняемую форму) из аминокислот на мРНК = многие составляющие получаются из многих с участием относительно малого числа участников полимеразного процесса (хотя их число измеряется многими десятками – белков и РНК)
Многочисленные белки, которые синтезированы в полимеразных процессах с участием относительно малого числа составляющих, по сути, выполняют все перечисленные функции синтеза и другие функции
(!) здесь же заложены и степени свободы: если добавить в ДНК дополнительную длину, отвечающую еще каким-то генам или прообразам генов, то добавка (с переводом по коду в белок) будет воспроизводиться. При этом есть риск, что такие добавки плохо подействуют на воспроизводство исходной отлаженной совокупности (как «мусор» или хуже, вплоть до «яда») + за счет дополнительной нагрузки может не сойтись баланс скоростей получения дополненной совокупности и ее деградации
В результате полимеразные процессы за счет относительно немногих участников (непосредственных исполнителей) всех трех полимеразных процессов позволяют организовать получение мономеров для них самих
(!) формально можно представить, что при биологическом воспроизводстве происходят только три эти процесса как основная и единственная цель (если выполнение множества других функций рассматривать именно как их обеспечение – мономерами, условиями среды и т.п.; тогда получение других мономеров, например, для мембран и клеточных стенок является способом обеспечить эти три процесса)
Количество генов (и соответственно белков, 1 ген = 1 белок, но возможно, в различных модификациях) как характерный размер надстройки:
(О) эукариоты – одноклеточные и многоклеточные организмы с ядром (греч. «эу» – хорошо, полностью; «карио…» – принадлежность к клеточному ядру)
(О) прокариоты (доядерные) – только одноклеточные, не имеют ядра (как компартмента, ограниченного мембраной) и хромосомного аппарата
(Д) прокариоты – 3000 генов (кишечная палочка), эукариоты – 30000 генов (человек)
Эти значения приведены как грубый ориентир (среди эукариот есть и одноклеточные, соответственно число генов для них близко к кишечной палочке)
+ Для сравнения (неочевидность приведенных различий): размер генома отличается на 3 порядка (в парах нуклеотидов – п.н.): кишечная палочка 4,2 106 п.н., человек 6 109 п.н.
(!) все необходимы (без них воспроизводство невозможно) или существенны (много теряем – всегда или в какой-то важной ситуации)
с. 35 ННМ:
Метаболизм (обмен веществ) = ассимиляция + диссимиляция
Пластический обмен = ассимиляция
Энергетический обмен = диссимиляция
(М) это значит, что адекватное описание должно включать многие тысячи уравнений даже для простейшего живого организма.
Отсюда же обширная перспектива теоретического описания: вместо одной оптимизации в физике три оптимизации в биологии (физическая, как формальная типа принципа наименьшего действия + оптимизация как следствие текущей конкуренции + оптимизация как следствие перспективной/эволюционной конкуренции) + другие общие ограничения
= образ: «пирог» теоретического описания оказался настолько большим, что понятно, почему так долго не получалось освоить («отъесть хоть сколько-то большой кусок»)
Начинаем «есть» этот «пирог», по сути, уже надрезали по нескольким направлениям:
–организация схемы воспроизводства (в частности, ее сходимость) влечет много ограничений
–разделение типов процессов (воспроизводство как холостой ход, воспроизводство непредсказуемости, энергетика)
–три оптимизации – это разные характерные времена
Иными словами, уже обозначено разделение переменных, дальше будем добавлять еще направления разделения переменных (другие и внутри этих) и развивать эти
Не вошло в лекцию:
(П) схема воспроизводства в явном виде – воспроизводство бактериофага MS2 (параллельно после 3-й лекции на общем семинаре для 1-го курса)
Потенциал воспроизводства велик, поэтому общее число составляющих и соответственно превращений при их получении велико, начнем с одного этапа – отдельного физико-химического превращения
Образ: начнем с маленького «кусочка» огромной схемы
Напоминание: использовали язык молекулярной кинетики, он важен и интересен не только для биологии, но и для описания воспроизводства как размножения некоторых единиц (в экономике, применительно к некоторому производству и т.п.). В этом смысле есть естественная широкая перспектива: строим что-то сложное и интересно организованное, а затем «размножаем» некоторым способом
+ идея максимально возможного выражения однородности (для упрощения теоретического анализа) как универсального представления
схема со стрелками, которая была использована при обсуждении катализа (как механизма, обеспечивающего потенциал воспроизводства на молекулярном уровне), и является таким универсальным представлением
(К) это логично: если универсальное представление представляет все, то оно должно представлять и катализ
Рассматривали кинетические уравнения на примере химического превращения (заодно вспоминали, как писать такие уравнения)
A+2B → P (как было выше)
Для схемы (*) (с переобозначением констант скорости по номеру этапа, для обратных этапов – знак «–» в индексе):
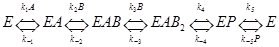 ,
,
d[E]/dt = – k1[E][A] + k–1[EA] + k5[EAB2] – k–5[E][P]
d[EA]/dt = k1[E][A] – k–1[EA] – k2[EA][B] + k–2[EAB]
d[EAB]/dt = …
d[EAB2]/dt = …
d[EP]/dt = …
Достаточное условие упрощения кинетического описания:
A0, B0, P0… >> E0 = [E]+…
(!) тогда есть теорема о возможности приближенно заменить все кинетические уравнения, кроме одного, на алгебраические уравнения
= утверждение о квазистационарности концентраций (КСК)
Простейшее обоснование применимости утверждения о квазистационарности концентраций (КСК) следует из оценки характерных значений производных при соотношении концентраций A0 >> E0.
Для субстрата (при максимальной нестационарности, когда за характерное время превращения τ концентрация субстрата уменьшается от A0 до нуля) производная dA/dt порядка ΔA/τ, т.е.
dA/dt ~ ΔA/τ = A0/τ
Аналогично
d[EA]/dt ~ Δ[EA]/τ < E0/τ
Значения производных можно представить в виде:
– d[A]/dt = v1 – v–1 = vin;
d[EA]/dt = (v1 – v–1) – (v2 – v–2) = vin – vout.
В силу d[A]/dt = vin << d[EA]/dt = vin – vout, как следует по оценке производных выше, vin приближенно равно vout. Аналогично можно обосновать приближенное равенство других пар скоростей v2 – v–2 и v3 – v–3 и т.д., в результате скорость превращения
v = v1 – v–1 = v2 – v–2 = v3 – v–3 = …
более подробное доказательство применимости утверждения о квазистационарности концентраций (КСК) – в ТБл
Факт: достаточное условие практически всегда выполнено, исключая несколько частных ситуаций (РДФК + еще несколько)
+ (!) для стадий порядка выше первого возможно представление эффективной константой скорости первого порядка (если включить в настоящую константу все концентрации, кроме той, что изменяется быстро). Например, для диффузионных этапов в эффективную константу скорости можно включить концентрацию субстрата или продукта, т.к. они изменяются гораздо медленнее, чем концентрации всех комплексов с белком – катализатором, переносчиком и т.д.
в данном случае: включение в эффективную константу скорости концентрации второго реагента для реакций 2-го порядка
(!)
= повышение фактической симметрии универсальной схемы (которая тогда включает только этапы 1-го порядка, является циклической и т.д.)
(М) это позволяет получать выражения для скорости через учет всей совокупности свойств, которым оно должно удовлетворять (см. ниже)
Но прежде, чем использовать схемы с повышенной симметрией, нужно разобраться с диффузионными этапами
Диффузионные этапы имеют энергию активации 8–12 кДж/моль (в несколько раз меньше суммы энергий слабых связей для молекулы в водной среде, для молекул воды в среднем 3,4 связи энергией 18,8 кДж/моль при 20оС) и поэтому константой обратного процесса
k–D = 1013 с–1 10–2 = 1011 с–1,
Отсюда для прямой стадии значение kD порядка 1010 М–1с–1. Это следует из геометрических соображений. Удобно взять концентрацию диффундирующих молекул A и B (например, субстрата и фермента) 1 М, для воды в любом случае 55 М (1000/18), т.е. в модели лототрона на 55 шаров воды приходится по одному шару молекул, которые должны оказаться рядом для взаимодействия. При координационном числе (число шаров в окружении, с которыми возможно взаимодействие), равном 6, значение kD численно примерно на порядок меньше, чем k–D (т.е. в модели одинаковых шариков k–D/kD ~ 10 М), в силу
v+ = kD[A][B] = v– = k–D[A…B].
При дополнительных ограничениях kD относительно уменьшается, как обычно бывает, т.к. реально имеем ситуацию, которая существенно отличается от обсуждаемого простейшего варианта одинаковых шариков трех типов. Участники взаимодействий, как правило, больше или существенно больше, чем молекулы воды, в частности, «шарик Е» (белок-катализатор или макромолекулярный комплекс) – это молекула, которая по массе и объему в 1000–10000 раз больше молекулы воды, а низкомолекулярные субстраты больше ее в несколько раз или на порядок. При взаимодействии субстратов с ферментами (конфигурация шарик – луза) характерное значение kD =108 М–1с–1 и меньше («Биокинетика», с. 77).
Следствие: характерные концентрации субстратов биохимических превращений как относительно низкие – порядка 10–5–10–6М (больше нет смысла, т.к. лимитирует химический этап, для которого k не больше 103 с–1) (М) оценка в приближении, что концентрации всех промежуточных комплексов порядка E0 + выражена необратимость (обоснование ниже)
|
|
|
|
Дата добавления: 2014-01-05; Просмотров: 211; Нарушение авторских прав?; Мы поможем в написании вашей работы!