КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Динамика популяций
|
|
|
|
Земля
Рис 3.2.
Величина непосредственного излучения, достигающего данной точки поверхности Земли колеблется в пределах 0,3 – 2,4 Дж/см2 мин и определяется рядом условий: толщиной и оптическими свойствами атмосферы, углом падения лучей, положением Солнца. Т.е. непосредственное излучение в определенном месте земного шара зависит от географической широты, времени года, времени суток и погодных условий.
Солнечное излучение в результате контакта с частицами атмосферы подвергается частично рассеиванию и отражению. Оно достигает поверхности Земли под разными углами. Благодаря этому, солнечная энергия проникает в места, закрытые для прямого солнечного облучения, а в пасмурные дни является единственным источником солнечного излучения. В течение суток величина рассеянного излучения возрастает до полудня, а затем уменьшается. Энергия рассеянного излучения зимой в два и более раз больше энергии (для Европы) непосредственного излучения и примерно равна ей в другие времена года. Распределение энергии рассеянного излучения проявляет сдвиг в сторону ультрафиолетового излучения, доля которого в нем выше, чем в непосредственном излучении Солнца. Отсюда понятно, почему можно загорать в тени при совершенно безоблачном небе.
Сумма непосредственного и рассеянного излучений представляет полное излучение Солнца. Часть этого излучения отражается и направляется обратно к небу, не меняя своей длины волны. Мерой отражения излучения является АЛЬБЕДО, которое определяет отношение (в %) отраженного излучения к падающему на данную поверхность. Величина альбедо зависит от характеристики поверхности. Альбедо сухого чистого снега может достигать 95 %, а влажной темной почвы - только 5 %. Значения альбедо зависят от высоты Солнца - лучи, падающие на поверхность воды под острым углом, могут отражаться на 90 %.
Часть полного излучения, поглощенного поверхностью Земли, превращается в тепловую энергию, вторично излученную в направлении неба в инфракрасном диапазоне. Излучение поверхности Земли в течение дня увеличивается с увеличением температуры почвы, а ночью уменьшается. Длинноволновое излучение Земли большей частью поглощается уже в нижней части тропосферы. Поглощение излучения поверхности Земли атмосферой приводит к увеличению излучения последней в том же инфракрасном диапазоне. Таким образом, значительная часть излучения Земли возвращается назад и теперь полностью поглощается. Напряженность длинноволнового излучения, возвращаемого атмосферой, оценивается в 2,5 Дж/см2 мин., а безвозвратные энергетические потери в космическое пространство, в зависимости от облачности, составляют 0,4 - 1,3 Дж/см2 мин. Т.е. обратное излучение атмосферы является эффективным способом возврата энергии, потерянной ранее Землей. Это явление, известное под названием ПАРНИКОВОГО ЭФФЕКТА, существенным образом влияет на тепловой баланс Земли. Средняя температура Земли равна 15 0 С, тогда как без возвратного излучения она должна составлять -23 0 С.
Таким образом, баланс на активной поверхности* выглядит следующим образом:
S = SC + SH + SA - S3 - SL, Дж/см2 мин, (3.1)
где
SС - непосредственное излучение Солнца;
SН - рассеянное излучение неба;
SА - обратное излучение атмосферы;
S3 - излучение Земли;
SL = L (Sc + Sн)/100 - отражение Земли.
На спектральный состав и напряженность солнечного излучения влияют также локальные факторы - прежде всего, состав воздуха. Дым может поглощать до 90 % солнечного излучения.
Б. Энергия излучения в момент поглощения каким-либо веществом превращается в тепловую. Механизм транспорта тепла различен у жидкостей, газов и твердых тел. В атмосфере теплообмен осуществляется через излучение, перемешивание потоков воздуха, а также через испарение и конденсацию водяного пара. В гидросфере перенос тепла происходит турбулентным способом, благодаря течениям, волнению, конвекцией тепла и испарением. В почве передача тепла происходит только за счет теплопроводности. Эти различия являются причиной того, что годовые колебания температуры охватывают всю тропосферу, в воде - слой до нескольких сот метров, в почве - слой не более 10 - 20 м.
В. Вода играет важную роль климатообразующего фактора. Полное количество воды, находящейся на Земле, постоянно. Баланс водных ресурсов строится на предположении, что все
количество воды, поступившее с атмосферными осадками, равно количеству воды, испаренной с поверхности почвы и воды.
Гидросфера производит 84 % водяного пара, находящегося в атмосферном круговороте. Осадки над поверхностью океанов меньше испарения (77 %). Различие выравнивается поверхностным стоком. Реки возвращают в моря и океаны около 7 % полного количества воды, находящегося в круговороте. Влажная зона материков дает 10 %, а сухая зона - 6 % от общего испарения Земли. Во влажной зоне выпадает 17 %, а в сухой - 6 % осадков всего земного шара.
Полное количество водяного пара, находящегося в данном объеме воздуха, измеряется теми же единицами, что и атмосферное давление и называется упругостью водяного пара.
Состояние насыщения соответствует наибольшей упругости, возможной при данной температуре (равновесие испарения и конденсации). Отношение фактической упругости пара е и упругости в состоянии насыщения Е называется относительной влажностью
V =(е / Е) 100, % (3.2)
Изменение относительной влажности обычно обратно суточному циклу изменения температуры. Во влажных климатах колебание влажности невелики, а в сухих - значительны.
 |
* Активная поверхность - граница между двумя элементами системы, в которой происходит теплообмен; у почвы - ее поверхность, у воды - ее зеркало.
В слое 0 - 0,5 м над поверхностью земли влажность воздуха быстро возрастает по мере приближения к поверхности почвы. Поэтому стандартная высота измерения влажности принята равной 2 м.
В почвенной среде вода встречается в разных видах.
Химически связанная вода является составной частью различных соединений, входящих в состав почвы. Она не принимает участия в водном балансе. Вода почвы в парообразном состоянии имеет относительную влажность, приближающуюся к 100 %. Вода, связанная молекулярными силами с частицами почвы (ионы водяного пара), малодоступна для растений. Капиллярная вода заполняет волосовидные канальцы (сечением менее 3 мм2) между частицами почвы и является главным источником воды для растений с поверхностной корневой системой. Гравитационная (свободная) вода находится под действием сил тяжести, заполняет некапиллярные неплотности, перемещается вглубь почвы, легко доступна для растений. Грунтовая вода образуется из гравитационной, достигшей водоупорного слоя и здесь накопившейся.
Г. Условия жизни наземных организмов определяются атмосферным давлением. Для водных организмов аналогичным фактором является гидростатическое давление.
Наибольшие значения атмосферного давления наблюдаются в понижениях рельефа (ниже уровня моря), где оно может достигать 800 мм рт. ст., тогда как у границы вечных снегов в горах давление падает до 300 мм рт. ст.. Вертикальный градиент давления в значительной степени подобен ходу температуры.
Значительно большие перепады давления в водной среде. Гидростатическое давление с погружением на 10 м увеличивается на 1 атм.(760 мм рт. ст.). При средней глубине океанов 3800 м гидростатическое давление вблизи дна в 380 раз превышает давление воздуха на уровне моря.
При рассмотрении вопросов связи организмов со средой экология учитывает прежде всего критерии выживания и размножения.
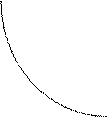
Условия среды проявляют значительную изменчивость во времени и пространстве. Поэтому большинство биологических видов приспособлены не к определенным значениям данного фактора, а к некоторым пределам его изменчивости. Как при более низких, так и при более высоких значениях фактора может наступить гибель организма. Наибольшее допустимое значение фактора называется верхней, а наименьшее - нижней критической точкой. Заключенный между этими двумя значениями диапазон изменчивости фактора представляет собой зону экологической толерантности. Вблизи критических точек активность организма сильно ограничена, тогда как середина зоны соответствует оптимуму (эта закономерность известна как «правило оптимума» (рис.3.3)).Толерантность различных организмов по отношению к одному и тому же фактору может существенно отличаться. У одних видов зона толерантности очень широка (такие виды называются эврибионтами), у других - узка (виды - стенобионты).
 2
2


 Интенсивность
Интенсивность
реакции
 организма
организма



1 3 Интенсивность фактора
1 - нижняя критическая точка
2 - точка оптимума
3 - верхняя критическая точка
Рис. 3.3.
Любое из условий существований, приближающееся к пределу толерантности или превышающее его, называется лимитирующим условием или лимитирующим фактором.
Рассмотренные ранее климатообразующие факторы являются лимитирующими. Приспособление организмов, адаптация их к тем или иным условиям существования применительно к климатообразующим факторам называется акклиматизацией. Акклиматизация происходит в течении ряда поколений и может закрепляться на генетическом уровне.
2. Адаптация к абиотическим факторам среды.
Для своего нормального развития и функционирования любой живой организм должен сохранять постоянство внутренней среды организма - ГОМЕОСТАЗ.
Изменение этого постоянства (гомеостаза) - ДЕЗАДАПТАЦИЯ - может привести к гибели организма. (Самая жесткая константа - кислотно-щелочное равновесие. Сдвиг рН на – 0,4 - гибель). Поэтому любой организм должен приспосабливаться (адаптировать) в условиях изменения экологических факторов. Под АДАПТАЦИЕЙ понимают совокупность морфологических *, физиологических, генетических и поведенческих приспособлений организма к определенным условиям среды.
Адаптация осуществляется при помощи нервной и эндокринной систем.
У живых организмов имеется три вида регуляции:
1. ГУМОРАЛЬНАЯ (жидкостная) - регуляция через жидкостную среду организма (кровь, лимфу, тканевую жидкость). Это самый древний вид регуляции. Он был характерен для древних организмов.
 * Морфология - наука о форме и строении организмов.
* Морфология - наука о форме и строении организмов.
Основные недостатки этого вида регуляции:
· замедленная реакция - регуляция происходит в течение определенного времени (например, при приеме таблеток, уколах);
· отсутствие конкретного адресата (оказывается воздействие на многие органы и системы организма с появлением побочных эффектов. Например, при приеме аспирина наблюдается внутрижелудочное кровоизлияние - 1 таблетка - 0,5 мл крови).
2. НЕРВНАЯ РЕГУЛЯЦИЯ. Появилась в ходе эволюции живых организмов. При этом типе регуляции воздействие фактора воспринимают рецепторы. (нервные окончания), передающие сигнал анализатором (группы рецепторов - органы чувств), и далее в центральную нервную систему (головной и спинной мозг), которая принимает решения и отдает исполнительную команду (ответная реакция).
Преимущества этого вида регуляции состоят в быстроте реакции (до 110 м/с), а также в воздействии на конкретный орган.
3. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ является разновидностью гуморальной регуляции. Осуществляется за счет гормонов, т.е. веществ, вырабатываемых в железах внутренней секреции. Это целенаправленный тип регуляции, но происходящий медленно.
Рассмотрим влияние абиотических физических (климатических) факторов среды на живые организмы и их приспособляемость (на примере адаптации к температуре).
В отношении солнечного излучения организмы стоят перед дилеммой: с одной стороны, прямое воздействие света на протоплазму смертельно для организма, с другой стороны - свет служит первичным источником энергии (об этом отдельная лекция о пищевых цепях), без которого не возможна жизнь. (Протоплазма - вещество клеток).
Ультрафиолетовое излучение. Высшие и низшие организмы по-разному реагируют на излучение этого диапазона. Весь участок ультрафиолетовых лучей губителен для бактерий. Уничтожающее действие усиливается при наличии в среде кислорода. Ультрафиолетовое излучение тормозит также развитие грибов и убивает их споры. Высшие растения в принципе не требуют УФ облучения для нормального развития.
Животные, имеющие крупные размеры тела, прежде всего, птицы и млекопитающие (в том числе и человек) нуждаются в некотором количестве УФ облучения в связи с синтезом витамина Д (отвечает за крепость костей скелета). Однако передозировка облучения кварцевой лампой у человека приводит к ожогам, а у мелких животных УФ облучение может вызвать гибель.
Видимое излучение. Является для организмов основным источником энергии и также летальным фактором среды. Его действие зависит от адаптации организма к свету.
Общее деление организмов с точки зрения их требований к напряженности видимого излучения подобно у растений и животных. Светолюбивые или световыносливые растения называются гелиофитами, животные - гелиофилами. Теневыносливые растения называются скиофитами; животные, избегающие света - гелиофобами.
Инфракрасное излучение. Зеленые растения пропускают или отражают большую часть лучей инфракрасного диапазона.
Животные используют инфракрасное излучение как источник тепловой энергии. Это излучение имеет большое экологическое значение, главным образом по причине вызываемого им теплового эффекта.
Адаптация к температуре. Жизнь может существовать в температурном интервале от -2000 С до +1000 С. Во льдах Антарктиды температура воздуха может понижаться до отметки -880 С, в африканских пустынях поднимается до +550С. На Земле есть регионы со значительными годовыми колебаниями температуры (например, на северо-востоке Украины до 600С), есть страны с незначительными (на Галапагосских островах круглый год температура воздуха примерно +270С). И во всех регионах Земного шара существует жизнь. Большинство видов и большая часть активности организмов приурочены к определенным, достаточно узким диапазонам температур.
Различают два типа живых организмов:
1. ПОЙКИЛОТЕРМНЫЕ - организмы, не имеющие постоянной температуры тела, т.е. температура их тела меняется в зависимости от температуры окружающей среды (холоднокровные животные - беспозвоночные, рыбы, земноводные и пресмыкающиеся).
2. ГОМОЙОТЕРМНЫЕ - организмы, не зависимо от температуры окружающей среды, поддерживающие температуру тела на одном уровне (теплокровные животные - птицы и большинство млекопитающих). Это становится возможным благодаря процессам ТЕРМОРЕГУЛЯЦИИ, происходящим в организмах.
Терморегуляция осуществляется путем изменения интенсивности теплообразования (при окислительных процессах в организмах) - ХИМИЧЕСКАЯ терморегуляция и путем изменения теплоотдачи через кожу (испарение пота и др.) - ФИЗИЧЕСКАЯ терморегуляция.
Температура тела в целом зависит от соотношения между количествами вырабатываемого и отдаваемого в окружающую среду тепла.
Единственным источником энергии в организме является пища, которая подвергается ряду обменных процессов, откладывается про запас, а затем, по мере необходимости, идет распад с выделением энергии. Наиболее интенсивно теплообразование в мышцах, печени, почках. Гораздо меньше - в соединительных тканях, костях, хрящах.
Наиболее интенсивные процессы тепловыделения происходят при сокращении мышц (примерно 80% энергии превращается в тепло, дрожь увеличивает теплообразование до 200 %).
В организмах животных на изменения температуры окружающей среды реагируют ТЕРМОРЕЦЕПТОРЫ, находящиеся в коже, на роговой оболочке глаз и в слизистых оболочках. Терморецепторы делятся на:
· воспринимающие холод (холодовые);
· воспринимающие тепло (тепловые);
Холодовые рецепторы залегают более поверхностно (на глубине 0,17 мм), тепловые - глубже (0,3 мм).
Количество точек на поверхности тела, где сосредоточены холодовые рецепторы примерно равна 250 тысячам, тепловые - примерно 30 тысячам.
В естественных условиях зимой преобладает химическая, а летом - физическая терморегуляция.
Реакция организма высших животных на низкие температуры происходит по следующей упрощенной схеме:
· первоначально низкая температура выступает возбуждающе, как стрессорный фактор;
· при дальнейшем воздействии температуры окружающей среды происходит снижение температуры тела и срабатывание механизмов сопротивления (начинаются процессы теплообразования - выделение энергии);
· при продолжении воздействия фактора на организм и исчерпании резервов последнего наступает гибель (у человека этому соответствует понижение температуры тела до +240 С, при которой сворачиваются ферменты).
При воздействии высокой температуры происходит выход крови из кровяных депо в кожу, расширение сосудов и увеличение интенсивности теплоизлучения.
Важную роль в отдаче тепла из организма играет процесс испарения влаги с поверхности кожи (через потовые железы) и легких. В течение суток потовые железы выделяют 500 мл влаги, с поверхности легких теряется 350 мл (всего 850 мл). Учитывая, что на испарение 1 мл воды требуется 0,58 ккал, можно увидеть, что таким путем организм может отдать в окружающую среду до 500 ккал в сутки.
У животных, не имеющих потовых желез, процессы испарения с кожи отсутствуют и теплоотдача идет с других поверхностей (например, у собаки - с поверхности языка, у зайца - с поверхности ушей).
Важным является отношение поверхности тела организма к его объему: чем оно меньше, тем меньше теплоотдача у животного. Поэтому у теплокровных животных, подверженных географической изменчивости, размеры тела особей статистически (в среднем) больше у популяций, живущих в более холодных частях ареала вида - ПРАВИЛО БЕРГМАНА.
Важным является также количество влаги в теле организма: чем ее меньше, тем легче переносится холод.
Влажность среды является важным фактором, определяющим возможность выживания наземных животных. По отношению к влажности организмы принято делить на 4 экологических группы:
1. Гидрофиты - организмы, живущие в воде.
2. Гелофиты - организмы, живущие на границе воды и суши.
3. Гигрофилы - организмы, требующие высокой влажности среды.
4. Ксерофилы - организмы, проявляющие высокую устойчивость к высыханию.
Атмосферное давление оказывает значительное влияние на ход жизненных процессов животных организмов. Для растений это влияние пока не установлено.
Испанские конкистадоры вынуждены были перенести столицу Перу в расположенную на берегу моря Лиму, т.к. на высоте 3500 м лошади, куры и свиньи не размножались. Оседлое проживание людей на высоте более 4000 м над уровнем моря приводит к вымиранию поселений.
Гидростатическое давление останавливает процессы брожения при 600 атм., а гнилостные процессы - при 700 атм. Рыбы поверхностных вод погибают при 300 атм., но известная всем морская звезда выдерживает давление до 600 атм.
Приспособляемость к новым климатическим условиям называется - АККЛИМАТИЗАЦИЕЙ.
Различают ИСТИННУЮ акклиматизацию, которая сопровождается изменением генетической структуры (происходит медленно и редко) и НАТУРАЛЬНУЮ, при которой происходят изменения в физиологии организма, но в пределах нормы.
Перечень лимитирующих факторов не ограничивается только физическими, абиотическими факторами (рис.3.1). Биологические взаимоотношения, биотические факторы не менее важны как регуляторы распределения и численности организмов в природе.
Понятие ПОПУЛЯЦИЯ является одним из центральных в экологии.
Одной из основных концепций ПОПУЛЯЦИОННОЙ экологии (ДЕМЭКОЛОГИЯ, изучает популяцию и ее среду обитания) является концепция САМОРЕГУЛЯЦИИ. Суть ее в том, что в популяциях существуют механизмы обратной связи, и воздействие внешней среды отражается в динамике популяции опосредованно, через эти механизмы. Данные механизмы весьма разнообразны и имеют различную природу: поведенческую, генетическую, физиологическую и т.д., однако в конечном счете они ведут к изменению демографических характеристик популяции. При этом отклик популяций на изменения зависит от внешних по отношению к популяции параметров среды.
В популяции существует внутривидовая конкуренция, т.е. борьба за ресурсы внутри вида.
Популяция обладает:
1. биологическими свойствами, присущими составляющим ее организмам,
2. групповыми свойствами, присущими только группе в целом. Такие свойства популяции, как плотность, рождаемость, смертность, возрастная структура и генетическая приспособленность характеризует популяцию в целом. Таким образом, особь рождается, умирает, но применительно к особи нельзя говорить о рождаемости, смертности, возрастной структуре.
Популяция имеет определенную организацию и структуру (временную и пространственную), которые можно описать.
Дадим определения и краткие описания основных свойств популяции.
Плотность популяции - это величина популяции в биомассе или количество особей, отнесенная к некоторой единице пространства. Плотности популяций млекопитающих охватывают диапазон почти в 5 порядков величин - от менее 0,001 до более 30 кг биомассы на гектар. Диапазон колебаний плотности популяций отдельно взятого вида намного меньше. Например, для медведя он составляет 0,1 - 1,0, а для хомяка - 0,12 - 1,2 кг/га.
То есть плотность, измеренная в биомассе, не зависит от размера особи.
Рождаемость - это способность популяции к увеличению численности.
Потенциальная или физиологическая рождаемость представляет собой теоретический максимум скорости образования новых особей в идеальных условиях. Она постоянна для данной популяции.
Реализованная или экологическая рождаемость обозначает увеличение численности популяции при фактических условиях среды. Она непостоянна, и может варьировать в зависимости от размера и возрастного состава популяций и физических условий среды.
Различают абсолютную рождаемость, определяемую путем деления числа вновь появившихся особей на время, и удельную - равную отношению абсолютной рождаемости к общему числу особей в популяции.
В 1988 г. в Украине на каждые 1000 человек рождалось и умирало в среднем 14,5 и 11,7 человека (для Харькова 13,8 и 10). Сейчас смертность превысила рождаемость.
Смертность отражает гибель особей в популяции. В определенной степени это понятие является антитезой рождаемости. Подобно рождаемости смертность можно выразить в виде абсолютной или удельной величины.
Реализованная или экологическая смертность - это гибель особей в данных условиях среды. Эта величина не постоянная и зависит от условий среды и состояния самой популяции.
Теоретическая минимальная смертность - величина постоянная для популяции, она представляет собой гибель особей в идеальных условиях, при которых популяция не подвергается лимитирующим воздействиям. Даже в самых лучших условиях особи умирают от старости. Этот возраст определяет физиологическую продолжительность жизни, которая всегда превышает ее среднюю экологическую величину.
Например, средняя продолжительность жизни, рождаемость, смертность существенно отличаются для государств с разным уровнем развития общества, т.е. с различными условиями существования.
Возрастная структура популяции является важной ее характеристикой, так как влияет на смертность и рождаемость. Соотношение разных возрастных групп в популяции определяет ее способность к размножению в данной момент и показывает, чего можно ожидать в будущем. В быстро растущих популяциях значительную долю составляют молодые особи. Популяции, находящиеся в стационарном состоянии, имеют более равномерное возрастное распределение. В популяции, численность которой снижается, будет содержаться большая доля старых особей, что и имеет место для ряда государств, в том числе для Украины.
На динамику популяций существенное влияние может оказывать половозрастная структура вида. Например, в зависимости от возрастной структуры существенно изменяется вероятность встречи брачного партнера, а следовательно, и рождаемость.
До сих пор мы говорили об изменениях в популяциях с течением времени. Однако популяции имеют и пространственную структуру.
Распределение особей популяции в пространстве может быть случайным, равномерным (более регулярным, чем при случайном распределении) и групповым (нерегулярным и неслучайным).
Случайное распределение наблюдается тогда, когда среда очень однородна и организмы не стремятся объединиться в группы.
Равномерное распределение встречается там, где между особями очень сильная конкуренция или существует антагонизм, способствующий поддержанию одинакового расстояния между особями. Чаще всего наблюдается образование различного рода скоплений.
Однако, если особи в популяции обладают тенденцией образовывать группы определенной величины, то распределение самих групп может оказаться более близким к случайному или даже равномерному.
В свою очередь пространственные структуры не являются чем-то застывшим и могут изменяться с течением времени, в том числе циклически или в виде волн плотности популяции в пространстве и времени. Примером может служить распространение вредителей в лесах и эпидемии. Для анализа этих явлений широко используются математический аппарат и моделирование на ЭВМ.
Поскольку популяционная динамика определяется взаимодействием рождаемости и смертности, то в идеале возможны два способа поддержания стабильной численности (когда рождаемость равна смертности):
1. Высокая смертность при высокой рождаемости (и соответственно малая средняя продолжительность жизни) - r - стратегия.
2. Низкая смертность при низкой рождаемости (большая продолжительность жизни) - k стратегия.
Наименования стратегий восходят к общепринятым обозначениям логистического уравнения популяционного роста Ферхюльста- Пирла, где под r понимается максимальная скорость роста численности популяции (репродуктивный потенциал), а под k - емкость среды, т.е. стабильная численность, к которой стремится популяция в данных условиях.
Согласно современным воззрениям r -стратеги имеют преимущество в нестабильных условиях, когда смертность мало зависит от плотности популяции, т.к. не пересыщенная организмами среда благоприятствует видам с высоким репродуктивным потенциалом (высоким отношением усилий на размножение к усилиям по поддержанию и выживанию особей).
k - стратеги лучше приспособлены к существованию в стабильных условиях, т.к. условия заполненной среды благоприятны организмам, характеризующимся низким потенциалом роста численности, но более высокой способностью конкурировать за скудные ресурсы и использовать их (больше энергии вкладывается в поддержание и выживание особей, чем в размножении).
Как правило, в природе любая из стратегий обладает преимуществом по сравнению с любым компромиссом.
Подробнее взаимодействие видов рассмотрим в следующем учебном в вопросе.
4. Взаимодействие популяций в сообществах
В сообществе существует межвидовая конкуренция, т.е. борьба за ресурсы внутри сообщества.
Существуют следующие типы взаимодействия двух популяций в сообществах:
1. НЕЙТРАЛИЗМ, при котором ассоциация двух популяций не сказывается ни на одной из них.
2. Взаимное КОНКУРЕНТНОЕ подавление, при котором обе популяции активно подавляют друг друга.
3. Конкуренция за ОБЩИЙ РЕСУРС, при котором каждая популяция косвенно отрицательно воздействует на другую в борьбе за функциональный ресурс. (Большинство видов существуют именно так, а не 2).
4. АМЕНСАЛИЗМ, при котором одна популяция подавляет другую, но сама не испытывает отрицательного воздействия.
5. ПАРАЗИТИЗМ (особи-паразиты меньше особей хозяина) и ХИЩНИЧЕСТВО (особи-хищники обычно крупнее особей-жертв), при которых одна популяция неблагоприятно воздействует на другую, нападая непосредственно на нее, но, тем не менее, сама зависит от объекта своего нападения.
6. КОММЕНСАЛИЗМ, при котором одна популяция извлекает пользу из объединения, а для другой это объединение безразлично.
7. ПРОТОКООПЕРАЦИЯ, при которой обе популяции получают от ассоциации выгоду, но эти отношения не обязательны.
8. МУТУАЛИЗМ, при котором связь популяций благоприятна для роста и выживания обеих, причем в естественных условиях ни одна из них не может существовать без другой.
Из указанных типов взаимодействий популяций наибольший практический интерес представляют конкуренция и хищничество.
Для выяснения действия различных факторов на изменение численности одного вида используют математические модели в форме дифференциальных уравнений скоростей роста популяции, на которую оказывает влияние другая популяция.
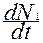 = r1
= r1 N1-
N1- 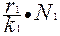 2 - α
2 - α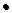 N1N2,
N1N2, (3.2)
(3.2)
где r - репродуктивный потенциал;
K - стабильная численность, к которой стремится популяция в данных условиях (емкость среды);
N1,N2 - численность популяций 1 и 2;
α- коэффициент, определяющий напряженность межвидовой конкуренции;
- скорость роста популяции 1;
r1N1 - максимальная скорость роста популяции 1;
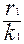 - определяет напряженность внутривидовой конкуренции.
- определяет напряженность внутривидовой конкуренции.
Из решений этих уравнений следует, что скорость роста каждой популяции равна максимальной скорости роста минус влияние собственной численности (напряженность внутривидовой конкуренции) и минус влияние другого вида (напряженность межвидовой конкуренции). Причем последний параметр (α) может быть положительным (см. типы взаимодействий двух популяций в сообществе п.п.2,3,4*,5), отрицательным (п.п.6*,7,8) и равным нулю (п.п.1,4*, 6 *), (где * для одной из двух взаимодействующих популяций).
При рассмотрении взаимодействия большего числа популяций уравнения составляют соответствующую систему уравнений.
Ограничиваясь взаимодействием двух видов, можно записать второе уравнение:
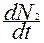 = r2N2 -
= r2N2 - 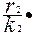 N22 - βN2N1, (3.3)
N22 - βN2N1, (3.3)
где β - коэффициент конкуренции, определяющий воздействие вида 1 на вид 2.
Система уравнений для двух популяций (3.2)-(3.3) носит название уравнений Лотки-Вольтерра, полученных ими независимо друг от друга в 1925 и 1926 г.г. и использованные В. Вольтерра для объяснения колебаний рыбных уловов в Адриатическом море. С тех пор эти уравнения были предметом многочисленных исследований.
Один из основных результатов состоит в том, что при конкуренции и отсутствии убежищ, один из видов с необходимостью вымирает, а другой достигает устойчивого состояния. И это соответствует как экспериментальным данным, так и реальному развитию природных экосистем.
В системе «хищник-жертва» картина совершенно иная. При отсутствии внутривидовой конкуренции (К1= К2= 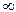 ) и конкурентного воздействия жертвы на хищника (β =0), а также наличия зависимости рождаемости (В 2) хищника от численности жертвы (В2 = Кn
) и конкурентного воздействия жертвы на хищника (β =0), а также наличия зависимости рождаемости (В 2) хищника от численности жертвы (В2 = Кn 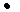 α N1 , где Кn - КПД переработки биомассы жертвы в биомассу хищника) получаем классическую модель В. Вольтерра
α N1 , где Кn - КПД переработки биомассы жертвы в биомассу хищника) получаем классическую модель В. Вольтерра
= r1N1- αN1N2
= (Кn α N1 - D2)N2, (3.4)
где B,D - функции рождаемости и смерти;
α N1 - скорость потребления жертвы одним хищником.
Анализ этой модели позволяет сделать вывод, что в системе «жертва - хищник» при соответствующих условиях (см. выше) возможен устойчивый режим колебаний численности жертв N1 и хищников N2.
Сплошная кривая на рис.3.4 называется предельным устойчивым циклом. Указано направление его прохождения (стрелки) и разметка по времени, например, в годах (цифры). Скорость прохождения цикла на данном участке определяется производной к кривой на этом же участке. В соответствии с устойчивым циклом происходят взаимосвязанные колебания численности популяций жертвы и хищника с одним периодом (в данном случае 12 лет). Такого рода зависимости были реально обнаружены. Они отвечают важнейшему свойству живой природы - самоорганизации. Действительно, этот цикл потому называется предельным, что к нему будут стремится все другие траектории соотношений численности хищников и жертв (пунктирные линии), независимо от их начального соотношения. Таким образом, хищник оказывает как бы стабилизирующее влияние на численность жертвы, не допускает ее взрывного увеличения и гибели в дальнейшем. Т.е., образно выражаясь, волки и лисы тоже нужны для устойчивого существования зайцев и косуль.
N1
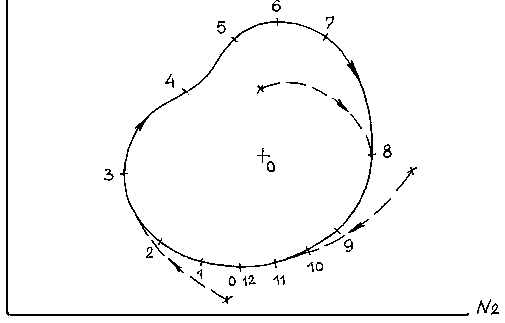
Рис. 3.4.
Данное решение является детерминистским, а реальная среда носит во многом случайный характер. В природе смертность и, особенно, рождаемость подвергаются случайным возмущениям. Причем случайные возмущения могут быть значительны по амплитуде и противонаправлены по действию на два взаимодействующих вида. В этом случае при неблагоприятных обстоятельствах, т.е. малой численности одного из видов, может произойти его вымирание. Такого рода ситуации неоднократно возникали и, увы, возникают в природе. Таким образом, чем меньше численность видов, больше размах противоположных случайных возмущений каждого из вида, тем скорее произойдет вымирание одного из них, т.е. разрушение сообщества.
Расчеты на моделях, учитывающих случайные возмущения, показали, что сообщество, состоящее из хищника и жертвы, не может стабилизироваться в случайной среде при низкой смертности хищника наряду с высокой доступностью жертвы или при высокой смертности хищника и труднодоступности жертвы. Такой вывод интуитивно понятен и соответствует реальным ситуациям в природе.
Следует также отметить, что применение математических моделей позволило сформулировать, количественно определить и оценить влияние на популяции и сообщества двух важнейших биотических факторов экологической среды - внутривидовой и межвидовой конкуренции.
Человек, к сожалению, еще очень медленно учится быть «расчетливым хищником», т.е. не истреблять свою жертву полностью при эксплуатации ее как ресурса. Теоретически оптимальное изъятие из эксплуатируемой популяции (например, рыбы из озера) должно происходить до уровня, при котором дальнейшая скорость роста популяции максимальна. Реально оптимальный выход живой продукции обеспечивается в объеме несколько меньшем половины предельной численности популяции при данных условиях среды обитания.
Вообще, в экологии свойства какой-либо биосистемы восстанавливаться после того, как ее структура и функции были нарушены, называется упругой устойчивостью биосистемы. Это понятие тесно связано со свойством живой материи к самоорганизации (рис.3.4). Способность же биосистемы сопротивляться, поддерживая свою структуру и функции неизменными, называют резистентной устойчивостью. Одна и та же биосистема обладает, как правило, только одним типом поддержания стабильности.
Еще одно исключительно важное понятие в биоценологии - экологическая ниша. Она включает в себя не только физическое пространство, но и определенную часть экологической среды, описываемой, как известно, абиотическими и биотическими факторами. Следовательно экологическая ниша данного вида определяется не только местом, где он живет, но и его требованиями к окружающей среде (температура, влажность и т.д.), а также взаимоотношениями с другими видами при добывании пищи. Конкуренция за один и тот же пищевой ресурс приводит к уменьшению ниши одного из видов или даже его полному вытеснению из данной ниши. Вытеснение одного вида другим может произойти и в результате конкуренции по любому другому параметру экологической ниши, например, количеству поступающей солнечной энергии (у растений).Зона экологической толерантности вида соответствует ширине экологической ниши по данному фактору. Для характеристики ниши используют еще ее перекрывание экологическими нишами соседних видов, без чего не обходятся реальные биоценозы. Виды,занимающие одинаковые ниши в разных географических областях, называются экологическими эквивалентами.
Давая определение биогеоценозу, как совокупности всех видов, населяющих определенную территорию, мы фактически не накладывали никаких ограничений на географические характеристики этой территории. Вместе с тем, границы природных биосистем не могут быть заданы произвольно. Поэтому введено понятие, уточняющее ранее используемый нами термин. Биогеоценозом называется совокупность популяций всех видов живых организмов, населяющих определенную географическую территорию, отличающуюся от соседних территорий по химическому составу почв, вод, а также по ряду физических показателей (высота над уровнем моря, величина солнечного облучения и т.д.). Т.е. пространство, занимаемое биогеоценозом, отделено существенным изменением на его границе хотя бы одного фактора экологической среды.
Разнообразие организмов на границе биогеоценозов имеет тенденцию к увеличению, что известно под названием краевого эффекта. Типичным примером может служить опушка леса, где разнообразие растений гораздо больше, чем в лесу и степи.
Биогеоценоз и его экологическая среда обитания вместе представляют собой биосистему самого высокого уровня организации (рис.3.1), называемую экологической системой или экосистемой. Понятие экосистемы часто используется и без ограничений на характеристики пространства и количество видов, его населяющих, т.е. может рассматриваться произвольное географическое пространство с произвольным числом видов.
Экосистема в более строгом понимании (имеющая в качестве биотического компонента биогеоценоз) обладает интересным свойством, называемым экологической сукцессией и состоящим в изменении во времени ее видовой структуры. В отсутствие внешних нарушающих процессов сукцессия представляет собой направленный предсказуемый процесс. Более или менее непрерывная смена видов во времени напоминает эстафету. В результате появляются последовательности сообществ, называемых сериями, также сменяющие друг друга в данном географическом пространстве. Для последней серии в этой цепи развития экосистемы используют термин - климакс. Он теоретически должен быть постоянным во времени и существовать до тех пор, пока его не нарушат сильные внешние возмущения.
Сукцессия, начинающаяся на участке, который прежде не был занят (поток застывшей лавы), называется первичной сукцессией. Сукцессия, начинающаяся на площади, с которой удалено прежнее сообщество (лесная вырубка), называется вторичной сукцессией.
На ранних стадиях сукцессии преобладает r - отбор, то есть успешнее вселяются виды с высокой скоростью размножения и роста. Напротив, для поздних стадий сукцессии характерен k - отбор, т.е. преобладают виды с низким потенциалом роста, но с более высокой способностью к выживаемости в условиях конкуренции. Типичный ряд вторичной сукцессии в предгорной области включает стадии: голое поле, степь, злаки и кустарники, сосновый лес и через 150 лет - климаксный дубовый лес.
|
|
|
|
|
Дата добавления: 2014-01-05; Просмотров: 739; Нарушение авторских прав?; Мы поможем в написании вашей работы!